 The Chromophore of GFP
The Chromophore of GFP
The Chromophore of GFP
The Chromophore of GFP
GFP is unique among fluorescent proteins in that its fluorophore is not
a seperately synthesized prostethic group but composed of modified amino
acid residues within the polypeptide chain.
Upon limited papain digestion of GFP, a hexapeptide with the same
spectral properties as
denatured GFP is released. The amino acid sequences of the hexapeptides from
Aequorea and Renilla are:
[3]
Aequorea: FSYGVQ
Renilla: FSYGDR
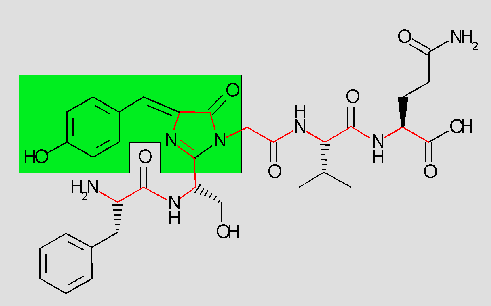
The chromophore itself is a p-hydroxybenzylidene-imidazolidone (green
background). It consists of residues 65-67 (Ser - dehydroTyr - Gly) of
the protein. The cyclized backbone of these residues forms the imidazolidone
ring. The peptide backbone trace is shown in red.
Although the amino acid sequence SYG can be found in a number of other proteins
as well, it is neither cyclized in any of these, nor is the Tyrosine oxidized,
nor are these proteins fluorescent.
This implies that the tendency to form such a chromophore is no intrinsic
property of this tripeptide.
Heim et al. [2] have proposed the following biosynthetic scheme for the chromophore. (R1 signifies residues 1-64, R2 residues 68-238.)
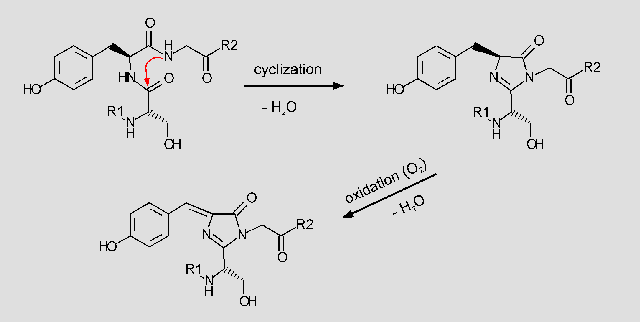
The first step is a nucleophilic attack of the amino group of Gly67 onto the carbonyl group of Ser65 (upper left). Subsequent elimination of water results in formation of the imidazolidinone ring (upper right). In a second step the bond of Tyr66 is oxidized to give a large delocalized pi-system (bottom). The proposed biosynthetic scheme is based upon the following observations:
Excitation and Fluorescence Emission Spectra of GFP
Denatured GFP is not fluorescent and its absorbtion spectrum is significantly different from native GFP [1], which implies that noncovalent interactions of the chromophore with its local environment have a great influence on the spectral characteristics and that fluorescence is mediated by amino acids close to the chromophore in the tertiary structure of GFP.
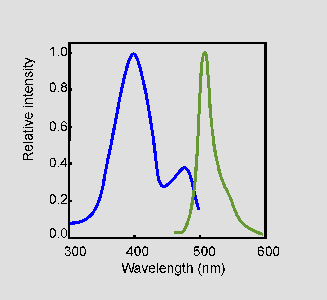
The excitation spectrum of native GFP from A. victoria (blue) has
two excitation maxima at 395 nm and at 470 nm. The fluorescence emission
spectrum (green) has a peak at 509 nm and a shoulder at 540 nm
[3].
(Spectra after [4])
Recombinant A. victoria-GFP generated in E. coli has the same spectral properties [4], the excitation spectrum of R. reniformis-GFP is different, whereas its emission spectrum is the same [3]. The absorption spectrum of A. victoria-GFP, is not significantly affected by titration of external pH up to pH 10, which is just below the threshold of denaturation [2, 11].
GFP is the first known example of a Förster cycle within the core of a protein [5,6,7]:
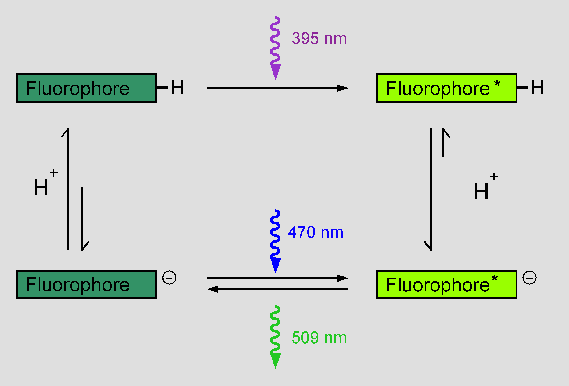
Depending on whether Tyr66 is in its hydroxyl form (upper left) or in its
phenolate form (lower left), the fluorophore absorbs light at 395 nm or at
470 nm, respectively. As phenols are known to be more acidic in their excited
state than in their ground state, it is postulated that the protonated excited
form of the fluorophore (upper right) converts to the excited phenolate (lower
right), which is the only fluorescent species and emits light at 509 nm.
As a result, a cycle is formed, in which the flourophore absorbs a photon, then
loses a proton, emits a photon and finally takes up a proton, returning to its
original state.
Interestingly, excitation at 395 nm leads to a decrease over time of the 395 nm
peak and a reciprocal increase of the 475 nm excitation band
[6].
After the structures of wild type GFP
[6]
and of the S65T variant of GFP [7] have
been solved by X-ray crystallography, some GFP variants, which have one
excitation band of wild type GFP deleted and retain the other without a significant
wavelength shift, can be understood in view of the 3-dimensional structure and
the Förster cycle:
The fluorophore inside the protein is not solvent accessible. It interacts with
the sidechains of many surrounding residues via hydrogen-bonds:
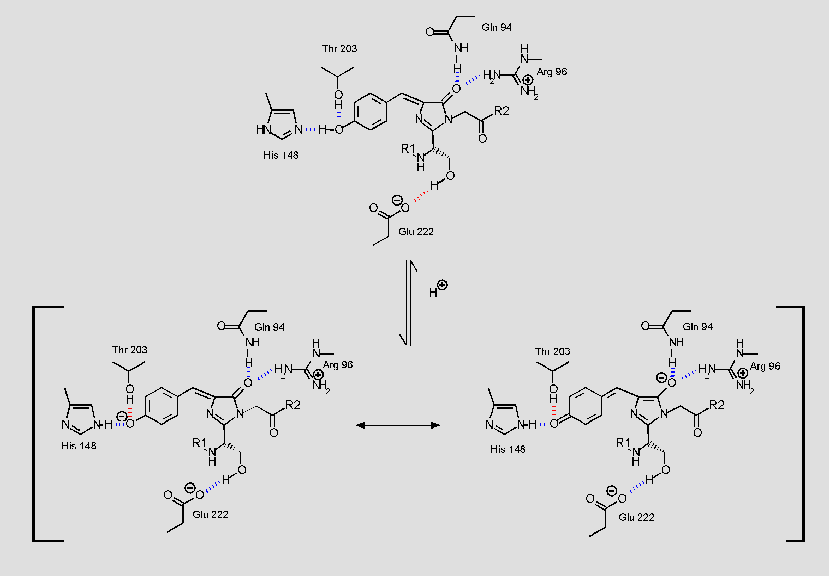 Click on the small picture for
a larger illustration (8k) of some of the fluorophore's interactions with the
surrounding sidechains. Hydrogen-bonds are shown in blue, except for the one
presumably stabilizing the respective form of the fluorophore, which is shown in
red.
Click on the small picture for
a larger illustration (8k) of some of the fluorophore's interactions with the
surrounding sidechains. Hydrogen-bonds are shown in blue, except for the one
presumably stabilizing the respective form of the fluorophore, which is shown in
red.
Stabilization of the phenolate or hydroxyl form of Tyr66 by specific hydrogen-bonds can explain why disruption of these hydrogen-bonds leads to an altered absorption spectrum in GFP-variants:
There is a vast amount of different GFP variants described in the literature, many of which have quite interesting characteristics. But discussion of all of these is far beyond the scope of this project and besides, there is no rational explanation yet for quite a few of the observed features. So I will limit further discussion of GFP variants to those which bear a mutation in the fluorophore itself.
![]() [Introduction]
[Introduction]
![]() [Index]
[Index]
![]() [GFP Structure]
[GFP Structure]
 Silke Jonda's PPS2 project
Silke Jonda's PPS2 project