(
 rasmol
image 300kb)
rasmol
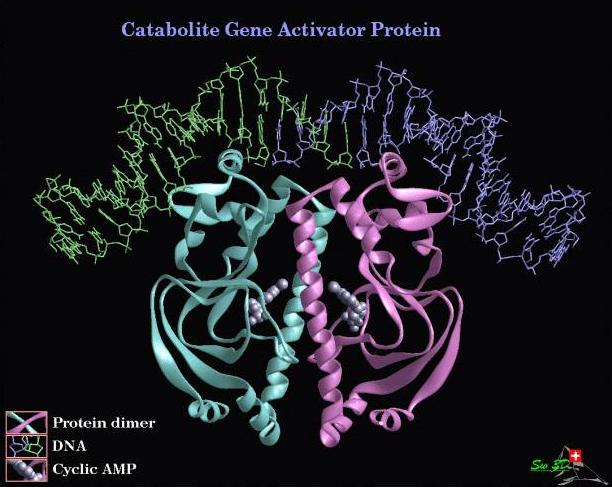
image 300kb)E.coli catabolite gene activator protein (CAP) in its active form is a homodimer of a 22.5 kDa protein requiring cAMP for binding activity. Its role in E.coli is in the regulation of genes involved in the uptake and catabolism of sugars. The DNA binding motif is helix-turn-helix. When bound to DNA this motif covers about three double helical turns of DNA (rasmol script). The recognition sequence is a 22 base pair palindromic sequence. A highly conserved TGTG / CACA sequence half a turn either way from the center of symmetry is contacted by a helix-turn-helix motif in the major groove. This penetrates parallel to the base pairs. The binding results in a ~40-45o kink in the DNA at each subunit (total 90o), closing of the major groove with concomitant widening of the minor groove. This bending of the DNA can be seen in the rasmol image above or in these images
 (54 kb jpg) ;
(54 kb jpg) ;  (67 kb jpg).
(67 kb jpg). The CAP protein is a regulatory protein and binds DNA without modifying it. The two example CAP-hybrids discussed below make use of the very large recognition sequence of the CAP protein to engineer rare cutting molecules. This large recognition sequence would mean the enzyme cuts less often (in 'random' DNA sequences) than the restriction enzymes readily available (recognition sites <=8 bp). This property could be of use in projects such as mapping of large genomes.
Both the molecules I discuss below were published by the same group in 1990 and 1994 respectively. (Ebright RH et al. 1990, Pendergrast PS et al. 1994)
CAP-1,10-phenanthroline
In this CAP derivative C178 of CAP (the only solvent accessible Cys) was converted into (N-Acetyl-5-amino-1,10- phenanthroline)-Cys. Cys178 is amino acid 10 of the helix-turn-helix motif and is brought in close contact with DNA upon CAP binding to DNA. This can be noted in the rasmol image of CAP (rasmol script). The agent that cleaves the DNA is the 1,10- phenanthroline-Cu(I) complex which is a chelating agent + metal, capable of indiscriminately cleaving DNA in reducing conditions.
It was found that this agent was capable site specific cleavage of DNA amongst sequence up to 7 kb in length however it was found that it had a tendency to cleave one strand only (75% of 40 bp substrate at least one strand and 35% both strands cleaved).
OP26CAP
For this CAP derivative a more elegant approach was taken. CAP was first modified by site directed mutagenesis C178 -> S178 and L26 -> C26, this left C26 the only solvent accessible Cys (Rasmol script). This Cys was then modified to yield [(((copper:o-phenanthrolin-5-yl)carbamoylmethyl)carbamoylmethyl)-Cys26;Ser178]CAP. This put the cleavage agent on amino acid 26 of both CAP subunits. It can be seen in the rasmol image of CAP that amino acid 26 of CAP is brought into contact with DNA only upon DNA kinking associated with site specific binding. This makes the agent very specific in its substrate requirements because while CAP can bind transiently to DNA with similar sequences (to its recognition sequence) it does not result in the large (40o) kink in the DNA. It was found that this agent was specific for cleavage of the CAP recognition sequence (in reducing conditions) in substrates up to 40 Mbp in size (reaction > = 70% completion) and in smaller substrates, 7.2 kbp the reaction went to > = 90% completion. The agent was found to cleave DNA at base 1 and 22 of the recognition sequence (on opposite strands) yielding a product with a 21 bp overhang. The positions of cleavage match well with the position of amino acid 26 in the crystal structure of CAP-DNA as seen in the rasmol image above.
This is a good example of protein design using two pre-characterized motifs and shows the potential of 'module' use in designing proteins.
In this example the Bzip (leucine zipper) DNA binding motif from Fos was coupled with the Cu(II), Ni(II) binding motif GKH producing a site specific DNA cleavage protein (Rasmol image of Fos139-198-Jun266-322
hetero-dimer (+/- one residue) bound to DNA in the two possible
orientations).Fos (an oncogenic transcription factor) binds to DNA as a hetero-dimer with Jun (another oncogenic transcription factor). Fos homo-dimers are not easily formed though Jun homo-dimers with DNA are readily formed (though these will not cleave DNA because the cleavage motif has been added to Fos). The consensus binding sequence for the Fos-Jun hetero-dimer is TGACTCA. This is not completely palindromic and depending on which way the hetero-dimer binds to DNA you get a slightly different kinking (see rasmol image).
The Cu(II), Ni(II) binding site motif GKH was introduced amino-terminally to the basic and leucine zipper regions of Fos (138-211). The resultant protein was capable of site specific DNA cleavage at the above consensus sequence. The hetero-dimer (GKH-Fos138-211 : Jun248-334) cleaved DNA at position 34 or 35 5' of the consensus sequence on the +ve sense strand (TGACTCA binding sequence) or 33 bp 5' on the -ve strand(TGAGTCA binding sequence) depending on the orientation of binding. Cleavage was relatively slow taking > 3 hours to go to completion even though the hetero-dimer was in excess to DNA.