Before entering into a discussion of stability in proteins, we must define exactly what we mean by stability. The word is used in different ways by different people. For example, a physical biochemist and a biotechnologist may each mean something different when they speak of stability.
The physical biochemist, on the one hand, would probably discuss protein stability primarily in terms of the thermodynamic stability of a protein that unfolds and refolds rapidly, reversibly, cooperatively, and with a simple, two-state mechanism:
![]()
Where Ku, is the equilibrium constant for unfolding.
The easiest proteins in which to study folding and stability are those that exhibit this sort of rapid reversibility. Both experimental design and also theoretical treatment of data are simplified by reversible systems. Thus, it is no surprise that most of the literature reports about stability discuss this type of reversible system. The bulk of this dissertation will also focus on thermodynamic stability.
In these cases, the stability of the protein is simply the difference
in Gibbs free energy, ![]() G, between the folded and the unfolded
states. The only factors affecting stability are the relative free energies
of the folded (Gf) and the unfolded (Gu) states. The
larger and more positive
G, between the folded and the unfolded
states. The only factors affecting stability are the relative free energies
of the folded (Gf) and the unfolded (Gu) states. The
larger and more positive ![]() Gu, the more stable is
the protein to denaturation.
Gu, the more stable is
the protein to denaturation.
![]()
The Gibbs free energy, G, is made up the two terms enthalpy (H) and entropy (S), related by the equation:
![]()
Where T is the temperature in Kelvin.
The folding free energy difference, ![]() Gu,
is typically small, of the order of 5- 15 kcal/mol for a globular protein
(compared to e.g. ~30 - 100 kcal/mol for a covalent bond).
Gu,
is typically small, of the order of 5- 15 kcal/mol for a globular protein
(compared to e.g. ~30 - 100 kcal/mol for a covalent bond).
The biotechnologist, on the other hand, is more concerned with the practical utility of the definition: Is the protein stable enough to function under harsh conditions of temperature or solvent? While the answer to this question may lie in thermodynamic stability (discussed above); it may also lie either simply in reversibility or, for irreversibly or slowly unfolding proteins, in kinetic stability.
If a protein unfolds reversibly it may be fully unfolded and inactive at high temperatures, but once it cools to room temperature, it will refold and fully recover activity. From a functional standpoint this may be all that is required for it to be classified as thermostable. However, from a thermodynamic standpoint (and in terms of this dissertation) it is classified as non-thermostable.
In the case of irreversible or slowly unfolding proteins, it is kinetic
stability or the rate of unfolding that is important. A protein that
is kinetically stable will unfold more slowly than a kinetically unstable
protein. In a kinetically stable protein, a large free energy barrier to
unfolding is required and the factors affecting stability are the relative
free energies of the folded (Gf) and the transition state (Gts)
for the first committed step on the unfolding pathway. Kinetic stability
is discussed in more detail in its own section; see Kinetic
Stability. Irreversible loss of protein folded structure is represented
by:
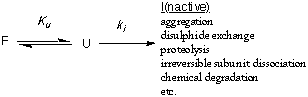
Where ki is the rate constant for some irreversible inactivation process.
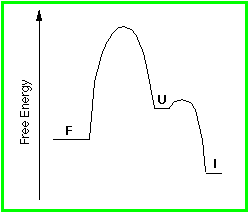
The free energy profile for a rapidly inactivating protein is shown below. Note that once the Unfolded form is reached, the energy barrier to inactivation is lower than that to refolding.