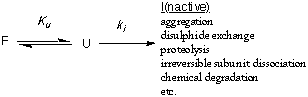
Kinetic stability, in the context discussed here, is a measure of how
rapidly a protein unfolds. It is a particularly important consideration
for proteins that unfold very slowly or denature irreversibly. A protein
can denature irreversibly if the unfolded protein rapidly undergoes some
permanent change such as aggregation (egg white) or proteolytic degradation
(subtilisin).
In cases such as these, it is not the free energy difference between the folded and unfolded state that is important. That will only effect the equilibrium, and it is not an equilibrium process. The important thing is the free energy difference between the folded and the transition states (activation energy), for it is the magnitude of this difference that determines the rate of unfolding (and hence inactivation).
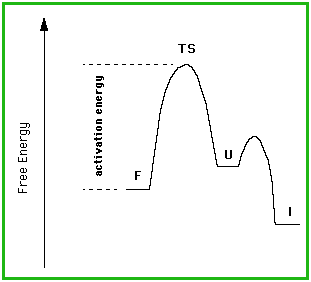
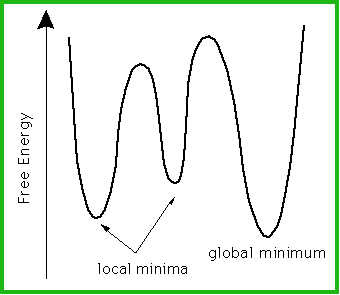
As an aside, this observation (in reverse: unfolded-to-folded) has some
consequences for the theory that a folded protein reflects the global energy
minimum for the structure. This is not true if the activation energy required
to reach that global minimum is so high that it cannot be reached. In such
a case, the structure will, to all intents and purposes, be trapped in a
local minimum, and that locally minimized structure will be the folded state
for that protein.
This case is illustrated by subtilisin, which requires a pro-sequence as a catalyst for folding; the pro-sequence is not required in the final structure, but lowers the activation energy of folding by more than 27 kcal/mol. In the absence of this pro-sequence, the low energy final folded structure would not be attained. This concept is discussed in more detail by Baker and Agard (1994).
As the folding / stability of few proteins that unfold irreversibly have
been studied in detail, I will discuss a mutational study on the kinetics
of unfolding of barnase disulphide mutants (Clarke
and Fersht, 1993). By means of SDM coupled with equilibrium and kinetic
folding and unfolding studies, the folding pathway and transition state
of this protein has been mapped (Serrano
et al., 1992 a & b).
 This picture illustrates
the folding of barnase.
This picture illustrates
the folding of barnase.
Disulphide bonds were introduced at positions that break early on the unfolding pathway (43 / 80), and at positions that break late on the unfolding pathway (85 / 102). The equilibrium stability and the unfolding kinetics were measured (Clarke and Fersht, 1993). (The stability was discussed in the previous section Disulphide Bonds.) The less stable 43/80 mutant unfolds 170 times more slowly, and the more stable 85/102 mutant only 17 times more slowly, when the disulphide bond is present than when it is reduced. Thus, the presence of an engineered disulphide linking regions that break early on the unfolding pathway gives rise to a 10-fold greater decrease in unfolding rate than the presence of disulphide linking regions that break late on the unfolding pathway; though the more rapidly unfolding mutant is more thermodynamically stabilized. The consequences of this for engineering or investigating kinetic stability in irreversible systems is that changes made in parts of the protein that are still fully formed in the transition state will have little effect on the observed rate of inactivation. Only changes to areas that have different structures in the folded and transition state will have an effect on the observed inactivation rate. Unfortunately, in the absence of the massive body of data available for barnase, it is not possible to predict which regions break early or late on the unfolding pathway, and trial and error is probably the best approach available.
For proteins such as barnase, in which even the most slowly unfolding disulphide mutant has an unfolding rate constant of approximately 1 mS, this analysis is not important to useful function. However, for proteins which unfold slowly and irreversibly, application of this principle is key. The hyper-thermostable alpha amylase from Bacillus lichiniformis unfolds slowly and irreversibly; unfolding by urea denaturation has a half-time of hours (S.Miraglia, unpublished). Mutants have been constructed in various parts of the structure, and even drastic mutations clustered around one region have little or no effect on stability (A. Day unpublished), suggesting that these regions are still fully formed in the transition state for unfolding.
![]() Disulphide Bonds
Disulphide Bonds ![]() Chemical Degradation
Chemical Degradation![]() Beginning
Beginning