The continuing health of an animal depends upon its ability to recognise and repel disease; this ability is called immunity.
Innate immunity , a first line of defence, is furnished by barriers such as skin, tears, saliva, and mucus, and the tissue inflammation that occurs after injury or infection.
Adaptive immunity develops specific defences against an invader that can be invoked whenever this particular intruder attacks again.
The immune system responds to surface structures of the invading organism called antigens. There are two types of adaptive immune responses: humoral and cell mediated.
In humoral immune responses antibodies appear in the body fluids and stick to and destroy antigens. The response is to toxic substances outside of the cell.
In the cell-mediated immune response cells that can destroy other cells become active (T-cells). They destroy disease infected cells or cells making mutant forms of normal molecules.
When disease associated proteins occur in a cell they are broken into pieces by the cells proteolytic machinery. Cell proteins become attached to antigen fragments and transport them to the surface of the cell, where they are "presented" to the bodies defence mechanisms.
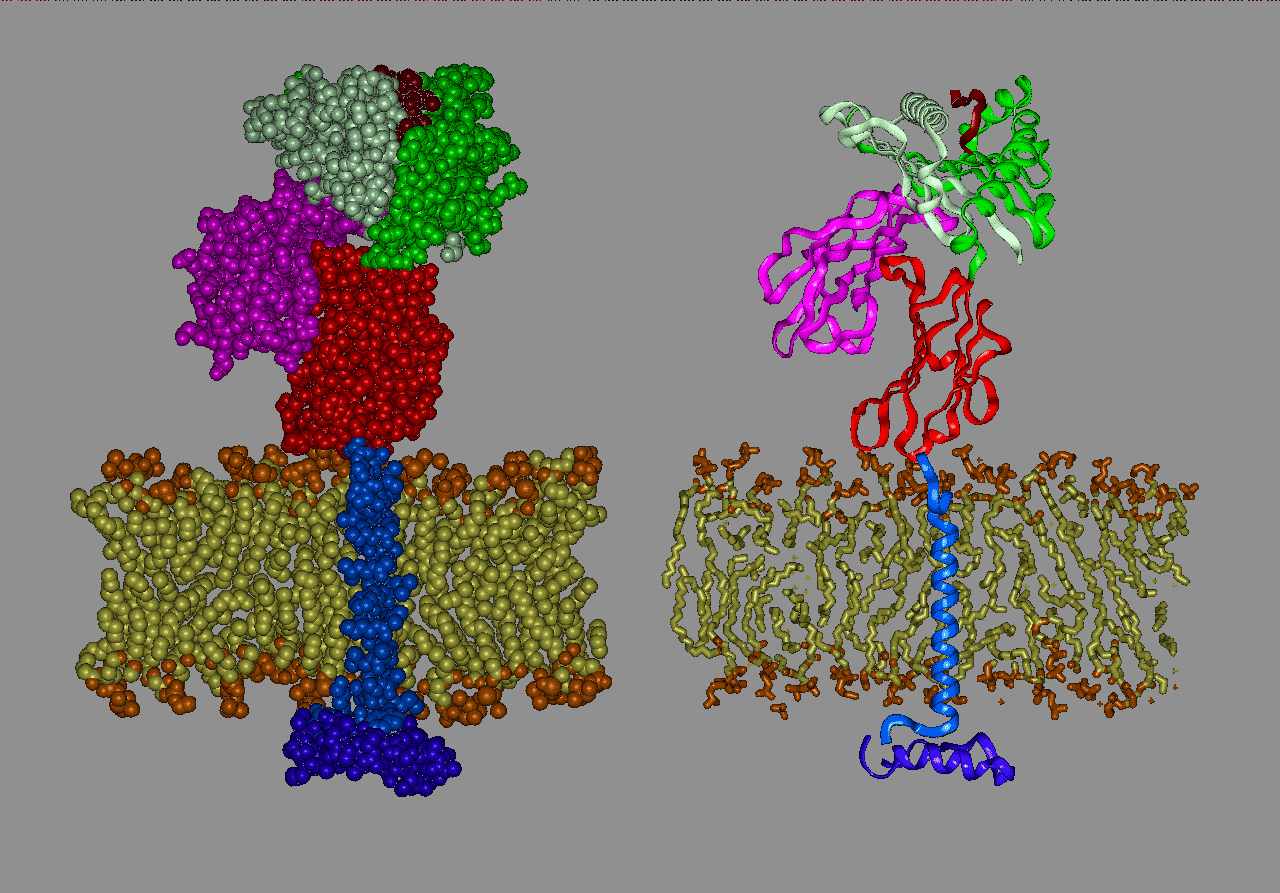
These transport molecules are called the Major Histocompatibility Complex (MHC) proteins. Without these, there would be no presentation of internal or external antigens to the T cells. The importance of MHC proteins is that they allow T cells to distinguish self from non-self. In every cell in your body, antigens are constantly broken up and presented to passing T cells. Without this presentation, other aspects of the immune response cannot occur.
Class I MHC proteins (found on all nucleated cell surfaces) present antigens to cytotoxic T lymphocytes (CTLs) . Most CTLs possess both T-cell receptors (TCR) and CD8 molecules On their surfaces. These TCRs are able to recognize peptides when they are expressed in complexes with MHC Class I molecules. For the TCR to bind a peptide-MHC complex two conditions must be met. Firstly, the TCR must have a structure which allows it to bind the peptide-MHC complex. Secondly, the accessory molecule CD8, must bind to the alpha-3 domain of the MHC Class I molecule. Due to genetic recombination events each CTL expresses a unique TCR which only binds a specific MHC-peptide complex. CTLs which recognize self-peptides (i.e. peptides produced by the normal host body as opposed to a foreign or cancerous cells) are removed in the thymus or tolerized after their release from the thymus. So, if a CTL can bind to a MHC-peptide complex on the cell surface, that cell is producing a peptide which is not native to the host.
The MHC Class II proteins (found only on B lymphocytes, macrophages, and other cells that present antigens to T cells), which primarily present peptides which have been digested from external sources, are needed for T-cell communication with B-cells and macrophages. Class II MHC proteins presenting antigens are detected by a different group of T cells (called T-helper or TH cells) to Class I MHC proteins (which are detected by CTLs cells).
The MHC proteins, and several closely associated with them in the carrying out of their functions, are coded for by loci that are close together within the Human Genome. Major Histocompatibility Complex proteins and their associated molecules are fundamental in the process of antigen presentation. The following pages collect and collate some of the information relating to this process.
Hughes AL, 1997
Molecular evolution of the vertebrate immune system.
Bioessays 19(9), 777-786 (1997)
EpiMatrix Tutorial
The Role of the Class I and II Major Histocompatibility Complexes.
Thompson MIP, Lecture
Major Histocompatibility Complex. James J. Thompson , Ph.D.
Histo at Birkbeck
A truly remarkable Web page.
This page was constructed as part of The Principles of Protein Structure '97
Accredited Advanced Certificate Course.
Please send mail relating to this page to:
John.Coadwell@bbsrc.ac.uk