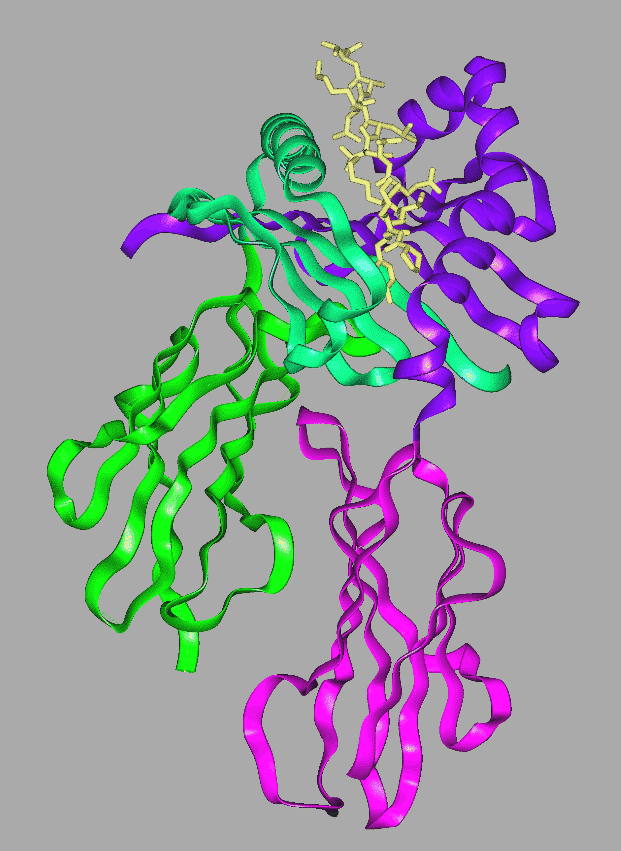
Although similar to Class I , the MHC Class II molecule is composed of two membrane spanning proteins. Each chain is approximately 30 kilodaltons in size, and made of two globular domains as shown in the ribbon diagram. The domains are named Alpha-1 (blue-green), Alpha-2 (green), Beta-1 (purple) and Beta-2 (magenta). The two regions farthest from the membrane are alpha-1 and beta-1. The two chains associate without covalent bonds. The bound peptide (cream) sits within the groove.
The three-dimensional structure of the Class II Human Histocompatibility Protein, HLA-DR1 has been determined at 2.8 angstroms resolution. If your Web Browser has been suitably configured, download this crystal structure of Human MHC Class II, [1DLH] (552Kb) - this is a dimer [Bbk|BNL|Hal]
The MHC molecules ability to present a wide range of antigenic peptides for T cell recognition requires a compromise between broad specificity and high affinity. Three-dimensional structures of both class I and class II MHC molecules show a unique structural solution to this problem. The peptide main chain is tightly bound whilst peptide side chains show less restrictive interactions. It is primarily the peptide side-chain contacts and conformational variability that ensures that the peptide-MHC complex presents an antigenically unique surface to T cell receptors.
Class II molecules are dimers consisting of an alpha and beta polypeptide chain. Each chain contains an immunoglobulin like region, next to the cell membrane. The antigen binding cleft, composed of two alpha-helices above a beta-pleated sheet, specifically binds short peptides, about 15 to 24 residues long. The amino acid sequence around the binding site, which specifies the antigen binding properties, is the most variable site in the MHC molecule . Differences between Class I and Class II structures can explain the different length requirements for the bound peptide. The ends of the antigen binding cleft of Class I molecules taper and are blocked by bulky tyrosines that bind the N terminus of the peptide. These conserved residues are not found in Class II molecules where smaller residues (glycine or valine) replace the larger tyrosines.
Broad specificity is accomplished through five cavities that accommodate the residue side chains of the bound peptide. The residues that line these pockets are highly polymorphic . This polymorphism contributes to the specificity of MHC Class II alleles. These pockets differ in the size and hypophobicity of the side chain they are able to accommodate. Because the main-chain carbonyl and amide groups are not symmetrically distributed relative to the side chains, there is a clear polarity imposed in the orientation of the bound peptide.
Castellino F, 1997
Antigen presentation by MHC class II molecules: invariant chain function,
protein trafficking, and the molecular basis of diverse determinant capture.
Hum Immunol 54(2), 159-169 (1997)
OMIM *142855
Lundberg AS, 1992
Evolution of major histocompatibility complex class II allelic diversity:
direct descent in mice and humans.
Proc Natl Acad Sci U S A 89(14), 6545-6549 (1992)