4.2 Identification without 3D Structure
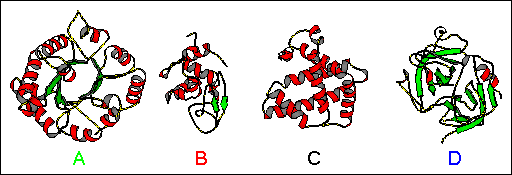
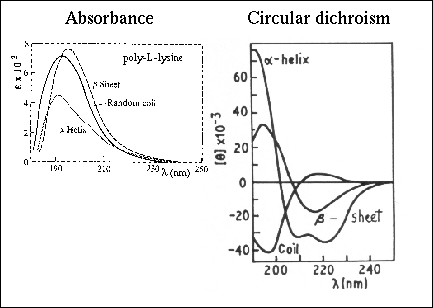
The phenomenon of circular dichroism is very sensitive to the secondary structure of polypeptides and proteins (Figure 21 and Figure 22 ). Circular dichroism (CD) spectroscopy is a form of light absorption spectroscopy that measures the difference in absorbance of right- and left-circularly polarized light (rather than the commonly used absorbance of isotropic light) by a substance. It has been shown that CD spectra between 260 and approximately 180 nm can be analyzed for the different secondary structural types: alpha helix, parallel and antiparallel beta sheet, turn, and other. A number of excellent review articles are available describing the technique and its application (Woody, 1985;Johnson, 1990). In fact, optical rotary dispersion (ORD, see below) data suggested a right-handed helical conformation as a major protein structural element before the Pauling and Corey model (Pauling & Corey, 1951) and Kendrew's structure of myoglobin. Modern secondary structure determination by CD are reported to achieve accuracies of 0.97 for helices, 0.75 for beta sheet, 0.50 for turns, and 0.89 for other structure types (Manavalan & Johnson, 1987).
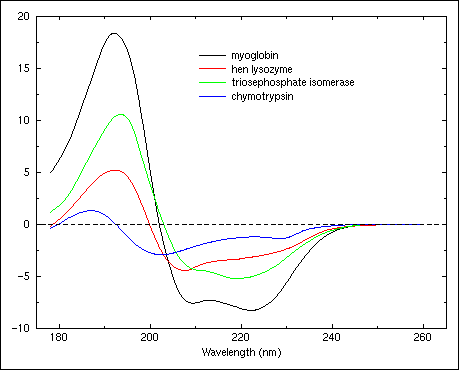
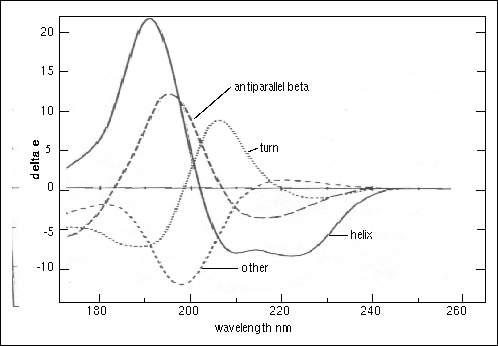
Figure 23. Circular dichroism spectra of "pure" secondary structures. Redrawn from Brahms & Brahms, 1980.
It can be shown that if right- and left-circularly polarized light is absorbed to different extents at any wavelength, there will be a difference in refractive indices at virtually all wavelengths. This accounts for the ability of small saturated chiral molecules to rotate the plane of polarized light of the D-line of sodium (589 nm), far away from absorptive bands. The dispersive nature of ORD is also the reason CD is a more sensitive analytical technique. Unlike the dispersive ORD phenomenon, circular dichroism can only occur within a normal absorption band and thus requires either an inherently asymmetric chromophore (uncommon) or a symmetric one in an asymmetric environment.
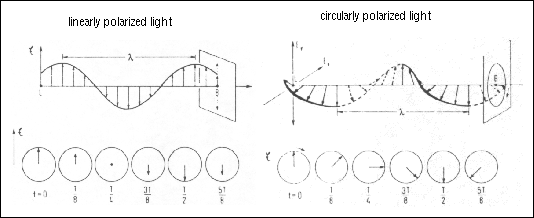
Most people are familiar with the concept of linearly-polarized light due to commonly-used Polarized sunglasses. However, circularly-polarized light may not be so easily conceived. Recall that electromagnetic radiation is a complex wave form that can be considered to behave as two simple wave motions at right angles to each other. One of these waves is magnetic (M) and the other is electric (E). Electromagnetic waves are generated by oscillating electric or magnetic dipoles and are propagated at the speed of light (c). Since the E- and the M-components are always perpendicular to each other, it is sufficient, in many cases, to consider only the E-component in describing the wave.
Figure 24. Schematic diagram showing the orthogonal electronic (E) and magnetic (B) components of linearly polarized light.
Although the amplitude of the E-wave oscillates in the zx-plane in the figure above, it could oscillate in any direction perpendicular to the direction of propagation (z). Unpolarized light, the type we get from the sun or a light bulb contains oscillations of the E-components in all directions perpendicular to the direction of propagation. Linearly polarized light results when the direction of the E-component is restricted to a plane perpendicular to the direction of propagation while its magnitude oscillates. Circularly polarized light is another form of polarization - in this case, the magnitude of the oscillation is constant and the direction oscillates.
Figure 25. Schematic diagram showing the electronic component of linearly polarized light (left) and right-handed circularly polarized light (right). Below each are the electronic component vectors as viewed along the axis of propagation from left of the diagram above.
The differential absorption of radiation polarized in two directions as function of frequency is called dichroism. When applied to plane polarized light, this is called linear dichroism; for circularly polarized light, circular dichroism. We can think of linear polarized light as the result of two equal amplitudes of opposite circular polarization. After passing through an optically active sample, circularly polarized light will be changed in two aspects. The two components are still circularly-polarized, but the magnitudes of the counter-rotating E-components will no longer be equal as the molar extinction coefficients for right- and left-polarized light are unequal. The direction of the E-vector no longer traces a circle - instead it traces an ellipse (actually an elliptical screw if we do not confine ourselves to the projection) There will also be a rotation of the major axis of the ellipse due to differences in refractive indices.
Historically, ellipticity is the unit of circular dichroism and is defined as the tangent of the ratio of the minor to major elliptical axis. Axial ratios of 1:100 will then result in an ellipticity of 0.57 degrees. Modern CD instruments are capable of millidegree precision. The unit ellipticity persists despite the fact that CD is now measured as the difference in absorbance of right- and left- circularly polarized light as a function of wavelength. Each type obeys the Lambert-Beer law so that the difference in extinction coefficients (left - right) at a given wavelength is equal to the difference in absorbance divided by the product of the pathlength and the concentration (Sorry, but I cannot format equations in html).
There exists a number of ways the CD of a sample has been reported in the literature. The most commonly used units in current literature are mean residue ellipticity (degree cm2 dmol -1) and the difference in molar extinction coefficients called the molar circular dichroism or delta epsilon (liter mol-1 cm-1). The molar ellipticity [theta] is related to the difference in extinction coefficients by [theta] = 3298(delta epsilon). Here [theta] has the standard units of degrees cm2 dmol -1 and the molar ellipticity has the units degrees deciliters mol-1 decimeter-1.
For proteins we will be mainly concerned with absorption in the ultraviolet region of the spectrum from the peptide bonds (symmetric chromophores) and amino acid sidechains in proteins in this exercise. Protein chromaphores can be divided into three classes: the peptide bond; the amino acid sidechains, and any prosthetic groups. The lowest energy transition in the peptide chromophore is an n --> p* transition observed at 210-220 nm with very weak intensity (emax xbb 100). Compare the UV and CD spectra of poly-L-lysine which can adopt three different secondary structure forms as a function of pH. The n --> p* transition appears in the a-helical form of the polymer as a small shoulder near 220 nm on the tail of a much stronger absorption band centered at 190 nm. This intense band, responsible for the majority of the peptide bond absorbance is a p --> p* transition (emax xbb 7000).
Figure 26. Comparison of the UV absorbance and the circular dichroism of poly-L-lysine in different secondary structure conformations. The figure on the left was taken from Cantor & Schimmel, 1980 and the figure on the right was taken from Campbell & Dwek, 1984.
Using CD, these different transitions are more clearly evident (no one has had success predicting secondary structure content from simple UV spectra). Exciton splitting of the p --> p* transition results in the negative band at 208 and positive band at 192.
The three aromatic side chains that occur in proteins (phenyl group of Phe, phenolic group of Tyr, and indole group of Trp) also have absorption bands in the ultraviolet spectrum. However in proteins, the contributions to the CD spectra in the near UV (where secondary structural information is located) is usually negligible. The disulfide group is an inherently asymmetric chromophore as it prefers a gauche conformation with css = xb1 90xb0 , corresponding to a right- or left-handed helical sense and can lead to a broad CD absorption around 250 nm. Aromatic residues, if unusually abundant, can have significant effects on the CD spectra in the region < 230 nm complicating analysis.
Like the other forms of absorption spectroscopy (UV/Vis, IR, etc.), CD is particularly powerful in monitoring conformational changes. In the region of 230-178 nm, one expects to observe effects of backbone conformational changes while CD effects at longer wavelengths (>230 nm) should isolate aromatic chromophore contributions and being environment dependent should reflect more global, three-dimensional properties of the protein.
If one is considering using CD spectroscopy, you should definitely read one of the reviews by the preeminent experimental CD spectroscopist W. Curtis Johnson jr (e.g., Johnson 1988 or Johnson, 1990; Johnson, 1992). To give you an idea what you will need, I offer the following brief summary of experimental conditions.
The concentration of peptide needed for CD studies will depend on exactly what information is expected from the spectra. As a general rule of thumb, one requires that the total absorbance of the cell, buffer, and protein be between 0.4 and 1.0 (theoretically, 0.87 is optimal). This means for a 0.01 cm cell, 20-50 ul of a protein concentration of 0.2-0.5 mg/ml is needed to record spectra to 178 nm. To achieve adequate signal-to-noise (S/N is proportional to the square-root of the amount of time measuring the spectrum), a spectra for secondary structure determination (260-178 nm) will require 30-60 minutes to record (plus an equivalent amount of time for a baseline as every CD spectrometer I have seen is a single beam instrument). Another consideration is that oxygen absorbs strongly below about 200 nm so very extensive purging with pure (oxygen-free) nitrogen (>16 liter/min) is necessary for these measurements. A typical buffer used in CD experiments is 10mM phosphate, although low concentrations of Tris, perchlorate or borate is also acceptable. Potassium fluoride is preferred to NaCl for increasing the ionic strength as chloride ion has a strong UV absorbance at low wavelengths.
In response to these shortcomings, several methods have been developed which analyze the experimental CD spectra using a database of reference protein CD spectra containing known amounts of secondary structure (Provencher & Glöckner, 1981; Hennesey & Johnson, 1981; Manalavan & Johnson, 1987; Sreerama & Woody, 1994). These methods are in general more accurate and reliable than the novel approach outlined above.
In one method (Manalavan & Johnson, 1987), single value decomposition is used to create orthogonal CD basis vectors from CD spectra of proteins with known secondary structure. Using the statistical technique of variable selection, unimportant variables are removed from an underdetermined system of equations allowing the solution for the important ones. The great advantage of this technique (and others like it) is that one makes no assumptions on the form of the CD from the individual secondary structural elements. One needs only to be sure that the overall structural characteristics of the analyzed protein are represented in the set of reference spectra. In this way irregularities of secondary structure and length dependencies should be taken into account
Two very important requirements for successful secondary structure analysis using these techniques deserves further comment. One is that the CD spectra need to be recorded from about 260 nm to at least 184nm (and preferable 178 or below; Johnson, 1990) and the other is that an accurate protein concentration (< 10% error) is essential. The assumptions and limitations of these techniques are discussed in detail by Manning (1989).
Circular dichroism spectroscpy has also been used to determine the tertiary structure class of globular proteins. The method proposed by Venyaminov & Vassilenko (1994) claim 100% accuracy for predicting all alpha, alpha/beta, and denatured proteins; 85% for alpha + beta; and 75% for all beta proteins.
The CD spectra of peptides have been reported in a number of solvent systems. The helix-promoting characteristics of trifluoroethanol (TFE) and hexafluoroisopropanol (HFIP) are well-known. A number of authors have argued that these solvents mimic a particular characteristic of the in vivo system and therefore attach relevance to structural studies performed in these solvents. Of particular importance to this matter is a study by Waterhous & Johnson (1994) who demonstrated that peptide sequences could be induced to form alpha helices or beta strands in TFE and non-micellar SDS solutions, respectively, regardless of their secondary structure in the native protein. This result underscores the difficulties in assigning relevance to structural studies in peptides using CD spectroscopy.
Generated with CERN WebMaker
Physical principles of CD
Inherently asymmetric chromophores (uncommon) or symmetric chromophores in asymmetric environments will interact differently with right- and left-circularly polarized light resulting in two related phenomena. Circularly-polarized light rays will travel through an optically active medium with different velocities due to the different indices of refraction for right- and left-circularly polarized light called optical rotation or circular birefringence. The variation of optical rotation as a function of wavelength is called optical rotary dispersion (ORD). Right- and left-circularly polarized light will also be absorbed to different extents at some wavelengths due to differences in extinction coefficients for the two polarized rays called circular dichroism (CD). Optical rotary dispersion enables a chiral molecule to rotate the plane of polarized light. ORD spectra are dispersive (called a Cotton effect for a single band) whereas circular dichroism spectra are absorptive. The two phenomena are related by the so-called König-Kramers transforms.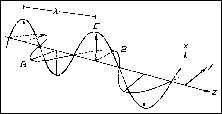
Secondary structure from CD spectra
The simplest method of extracting secondary structure content from CD data is to assume that a spectrum is a linear combination of CD spectra of each contributing secondary structure type (e.g., "pure" alpha helix, "pure" beta strand etc.) weighted by its abundance in the polypeptide conformation. The major drawback of this approach that there is no standard reference CD spectra for "pure" secondary structures. Synthetic homopolypeptides used to obtain reference spectra are in general, poor models for the secondary structures found in proteins. For example, the CD of an alpha helix has been shown to be length dependent and no homopolypeptide system has been found that is a good example of the beta sheet structure found in proteins. CD of peptides
Circular dichroism spectroscopy has been extensively applied to the structural characterization of peptides. The application of CD for conformational studies in peptides (like proteins) can be largely grouped into 1.) monitoring conformational changes (e.g., monomer-oligomer, substrate binding, denaturation, etc.) and 2.) estimation of secondary structural content (e.g., this peptide is 25% helical under these conditions). As already mentioned, CD is particularly well-suited to determine structural changes in both proteins and peptides. However, absolute structural content is more difficult and is prone to over-interpretation. Length-dependencies of the CD spectra of "pure" secondary structures are only one potential caveat.
No Title - 31 MAY 96
written by Kurt D. Berndt