After the processing in the Golgi, three types of N-linked oligosaccharides are formed :
The overall synthesis process is controlled by specific enzymes which act in the proper order and time, and are present in different amounts and in the respective Golgi compartment.
Each specific cell type in an organism contains its own specific processing enzymes. The result is that the same protein produced by individual cell types may have different carbohydrate chains.
For example certain Asn residues in HA (haemagglutinin) of influenza
virus have complex type oligosaccharides when the virus is grown in one
cell type but high-mannose oligosaccharides when the virus is grown in
another cell type.
The high-mannose oligosaccharides
have the same structures as the intermediates which are formed during the
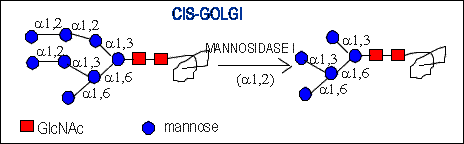
processing of complex oligosaccharides : (Man)8 (GlcNAc)2 , (Man)7 (GlcNAc)2
, (Man)6 (GlcNAc)2 , (Man)5 (GlcNAc)2. They occur after trimming by
Mannosidase I (alpha1,2 specific) in the cis-Golgi.
The exception is (Man)8 (GlcNAc)2 which is not cleaved by the
enzyme. Possibly the part of the oligosaccharide is buried in a creavasse
in the protein surface and thus is not available for mannosidase.
The complex type oligosaccharides
are formed after the further processing of (GlcNAc)2 (Man)5 by specific
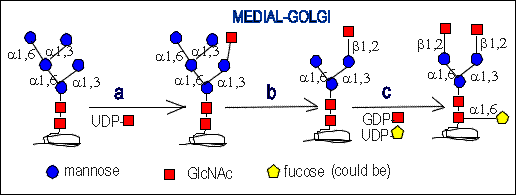
enzymes. This precursor is at first transported from cis- to the medial-Golgi
compartment. Then:
a.)N-Acetylglucosaminyltransferase I adds a GlcNAc residue to the
terminal mannose residue.
b.)Mannosidase II (alpha 1,3/6 specific) removes two mannose residues
c.)N-Acetylglucosaminyltransferase II adds one GlcNAc residue and
fucosyltransferase can add fucose residue but only when there is
GlcNAc linked (beta 1,2) to the mannose (alpha 1,3) branched.
Further processing occurs in the trans-Golgi :
d.)N-Acetylglucosaminyltransferase IV adds GlcNAc residue to
the mannose residue (then the oligosaccharide is higher branched). Higher
branched (tri-, tetra_ and pentaantennary ) complex type oligosaccharides
are formed by the subsequent addition of further GlcNAc residues by the
corresponding transferases to the mannose residues. The type is determined
by the order of action of the different N-Acetylglucosaminyltransferases.
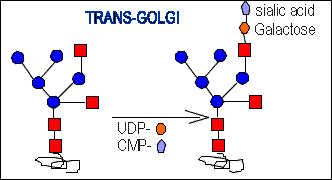
e.)Galactosyltransferase adds galactose residues to the terminal
GlcNAc residues.
f.)Sialyltransferase adds sialic acid residues which become the
terminal part of the oligosaccharide (they are thought to protect the glycoprotein
from degradation).
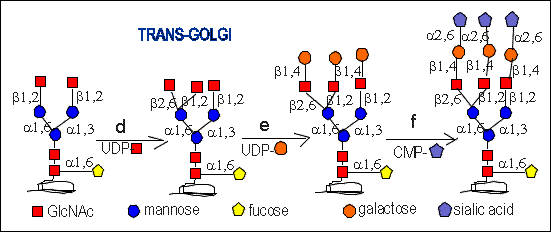
Sialic acid can be linked either (alpha 2,6) or (alpha 2,3) to the galactose
residues. It could be also linked (alpha 2,8/9) to another preceeding sialic
acid residue forming large sialic acid clusters (for example up to 55 in
N-CAMs - neural cells adhesion molecules). The sialic acid polymers are
found predominantly in neurons.
The hybrid type oligosaccharides are synthesized when the N-Acetylglucosaminyltransferase III acts prior to the action of Mannosidase II (in medial-Golgi compartment). GlcNAc-T III adds GlcNAc residue which is linked (beta 1,4) to the core mannose residue. It prevents the action of mannosidase II.
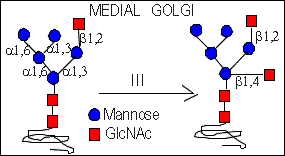
Then in the trans-Golgi galactose and sialic acid residues are added.
The carbohydrate chains of different types can undergo additional modifications such as sulphation, phosphorylation or acetylation.
Go to :
Biosynthesis of GPI membrane anchors