![]() Index to Course Material
Index to Course Material
![]() Index to Section 12
Index to Section 12
![]() Part 1;
Part 1;
![]() Part 2
Part 2
A simple reduction in the radius of the Fe ion is not sufficient to allow it to fit into the haem ring. Movement of the ion into the plane is prevented by steric repulsion between the proximal histidine side chain and the haem nitrogen atoms. This can only be avoided by a change in orientation of the histidine, which requires movement of the F helix to which it belongs. In fact the conformations of the turn between helices E and F, and the FG turn, also alter besides the shift in the position of helix F. The Tyrosine at the end of helix H (Tyr 140 in alpha subunit, Tyr 145 in beta) is expelled from the pocket between helices F and H, which leads to the breaking of the salt links between the chains.
The important interfacial structural changes are those between the two heterodimers (i.e. between the alpha 1 - beta 1 dimer, and the alpha 2 - beta 2 dimer), rather than between the two subunits within each of these dimers. Note that three of the four salt bridges listed in the section on the T state occur between the two heterodimers. Different hydrogens bonds stabilize the alpha 1 - beta 2 interface in the two different states (Exercise: identify these different hydrogen bonds) . There is a 15° rotation of the two heterodimers relative to each other, as indicated below.
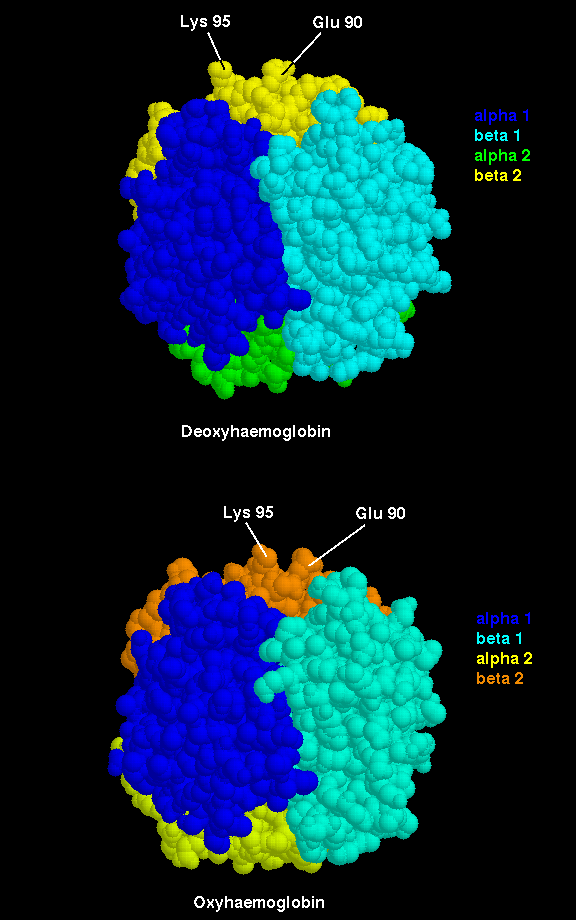 Click here for diagram.
Click here for diagram.
To obtain these views in the RasMol renditions of both the 1hho tetramer and the 2hhb structure, use the spacefilling rendition for the whole proteins and apply these commands to each:
reset rotate y -90 rotate z 45Alternatively, this SCRIPT will do everything, for either structure.
Therefore, binding of an oxygen molecule to the Fe ion in the haem ring leads to marked structural changes elsewhere in the molecule. The R-state tertiary structure of an oxygenated subunit stabilizes the same tertiary structure in the other chains, which therefore have a higher affinity for oxygen.
Because the key to cooperativity is the interaction between heterodimers, mutations in the interfacial residues between subunits alpha 1 and beta 1, and between alpha 2 and beta 2, do not tend to affect the cooperative binding system in the same way that mutations in the alpha 1 - beta 2 interface do.
The significance of the contacts between heterodimers also explains the observed increase in oxygen affinity. Suppose that a single subunit, say alpha 1, binds an oxygen and alters from the T to the R state. Bearing in mind the list of broken salt bridges listed previously, the interactions between (a) alpha 1 and alpha 2, and (b) alpha 1 and beta 2, will change. This would result in the R state tertiary structure in subunits alpha 2 and beta 2, so that the affinity of both for oxygen is increased. Indeed, the affinity for the third oxygen molecule is not markedly higher than that for the second, but both are much greater than that for the first. With oxygens now bound to alpha 1, alpha 2 and beta 2, the R state tertiary structure will also be stabilized in beta 1 (due to the tertiary structure of alpha 2 breaking the salt bridges between it and beta 1; refer to the previous list). The final empty chain will thus have a very stable R tertiary structure; this results in the dramatically higher affinity for the fourth oxygen.
Last updated 8th Jul '96