 Fig.
3.1.. Deca-glycine in PPII and PPI without hydrogen atoms, spacefill
modells, CPK colouring
Fig.
3.1.. Deca-glycine in PPII and PPI without hydrogen atoms, spacefill
modells, CPK colouring
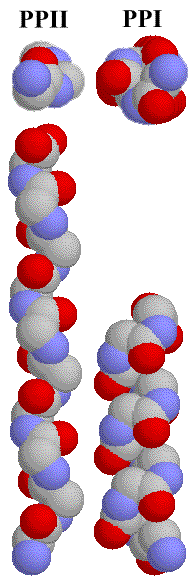
In Fig. 3.1. the picture of deca-glycine in PPII and PPI conformation is presented, without hydrogen atoms. From this, it can be clearly seen, that the more symmetrical PPII is a left-handed helix, with a higher rise, thus smaller diameter, than the right-handed PPI helix.
Fig.
3.1.. Deca-glycine in PPII and PPI without hydrogen atoms, spacefill
modells, CPK colouring
For comparison, here is a table of theoretical helical parameters of the PPII-, PPI-, and alpha-helix.
|
Type of helix |
Phi |
Psi |
Omega |
n |
r |
p |
H-bond |
| PPII |
-75 |
145 |
180 |
-3.0 |
3.1 |
9.3 |
- |
| PPI |
-75 |
160 |
0 |
+3.3 |
1.7 |
5.6 |
- |
| alpha |
-57.8 |
-47 |
180 |
+3.6 |
1.5 |
5.5 |
i,i+4 |
n is the number of residues per helical turn, positive value means
right-handed, negative means left-handed helix;
r is the helical rise per residue (Å), and p is the helical pitch
(Å/turn).
You can find the similar parameters and picture of other secondary structure types in Section 8 of PPS97.
As none of the poly-proline helices has an intra-chain H-bond, these secondary structure elements can not be identified by hydrogen-bond pattern characteristics, as regularly done for other types of secondary structure, but only by torsion angles. This may be one reason for having been assigned to the 'unordered' or 'random' group earlier.
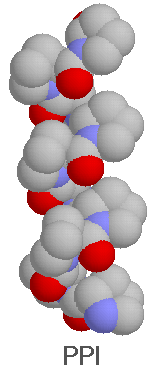 Fig.
3.2. Poly-L-proline in PPI conformation (PDB)
Fig.
3.2. Poly-L-proline in PPI conformation (PDB)
The formation of the right-handed PPI helix is possible only with proline residues because of the required cis conformation. There were no PPI helical segments identified (yet?) in proteins, but poly-L-proline in apolar solvents (alcohols with long carbon-chain) was found to adopt this conformation in some circumstances.
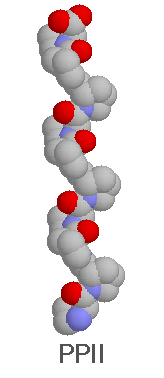 Fig.
3.3.1. Poly-L-proline in PPII conformation (PDB)
Fig.
3.3.1. Poly-L-proline in PPII conformation (PDB)
The PPII helix has much more biological importance.
It has been found in a large number of proteins, with little deviation
from the theoretical helix. The most important factor for distortion of
helices, is the possible cis conformation of proline. The length of PPII
segments is shorter (4-8 residues) than alpha-helical or beta-sheet
segments, which may be due to the lack of the stabilizing effect of intra-molecular
hydrogen-bonds. For the same reason PPII helices are more flexible than
alpha-helices. PPII helices has a rise per residue almost twice
as great as alpha-helices, so are much longer with the same number
of residues, and the main chain of PPII is more accessible. PPII helices
were found to be more exposed to solvent, than other secondary structures,
except beta-turns. PPII helices seem to be stabilized by main chain-water
hydrogen bonds (in the absence of main chain- main chain H-bonds), and
tend to have a regular pattern of hydrogen bonds with water. After this,
it is not surprising , that PPII helices are found mostly on the protein
surface, and has a preference, to hydrophilic residues (and proline). Pro
is the most preferred amino acid (and PPII is the most preferred structure
for Pro, so Pro residues can stabilize the structure), but any amino acid
can be found in a PPII helix, and helices were found without any proline.
Besides Pro, most often Gln, Ser, Arg and Ala are found in PPII regions,
Gly is rare.
The geometry of PPII helix allows this conformation to directly lead to,
or directly start from other secondary structures, and PPII helix can serve
as a bridge between different secondary structures. The PPII helix followed
by a beta-turn has been described as a novel superseconadry structure,
and has been suggested to to be the conformational requirement for proline
hydroxilation in nascent procollagen.
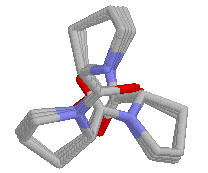
Fig. 3.2.2. Poly-L-proline in PPII conformation, viewed parallel
to the helix axis, presented as sticks, without H-atoms. (PDB)
It can be seen, that the PPII helix has a 3-fold symmetry, and every 4th
residue is in the same position (at a distance of 9.3 Å from each
other). This is important in when binding of PPII helices to SH3
domains.
Next: PPII helices in proteins
by Zoltan Szabo , 1997
{kind=link}
{kind=link}