 insulin, the growth factors,
glycoprotein hormones,
and the cytokines
(Glasgow &
Oxford).
insulin, the growth factors,
glycoprotein hormones,
and the cytokines
(Glasgow &
Oxford).
Back to Tertiary Structure Index
Individual domains may occur as `signalling'
proteins in the extracellular environment in higher organisms, more usually called hormones.
Examples include :-
insulin, the growth factors,
glycoprotein hormones,
and the cytokines
(Glasgow &
Oxford).
The tertiary structure of globular proteins reflects their interaction with their aqueous solvent. At a simple level, a globular protein may be considered to consist of a hydrophobic core surrounded by a hydrophilic external surface which interacts with water. The tertiary fold of the polypeptide is such that those residues with apolar side chains are buried in the centre, while the polar residues remain exposed. This principle is held by many to be the dominant driving force behind the folding of the polypeptide chain into the compact globular form: the aggregation and burial of the hydrophobic surface reduces the number of unfavourable interactions of these groups with water; thus the hydrophobic effect. (A section on this will soon be available as part of the "Overview of Molecular Forces" Chapter.)
Compare this distribution of hydrophilic and hydrophobic side chains with that in an integral membrane protein (see the previous section).
Bear in mind however that although the concept of "hydrophobic residues in, polar residues out" generally applies, it has its limitations. Residues with hydrophobic side chains have some polar surface area (the backbone atoms), and vice versa. There will necessarily be some buried polar groups in the hydrophobic interior (generally main chain atoms involved in hydrogen bonding in secondary structures). Likewise, the amount of solvent-exposed hydrophobic surface area is far more than minimal: in fact it constitutes approximately 50% of the total exposed surface area of a globular protein. A patchwork arrangement of hydrophobic and polar surface allows solvent molecules close to apolar regions to interact favourably with neighbouring polar surfaces.
Different types of globular proteins have been outlined in the Overview section of this chapter.
Another important technique for elucidating protein structure is Nuclear Magnetic Resonance (NMR) Spectroscopy (see also Imperial College's NMR stuff).
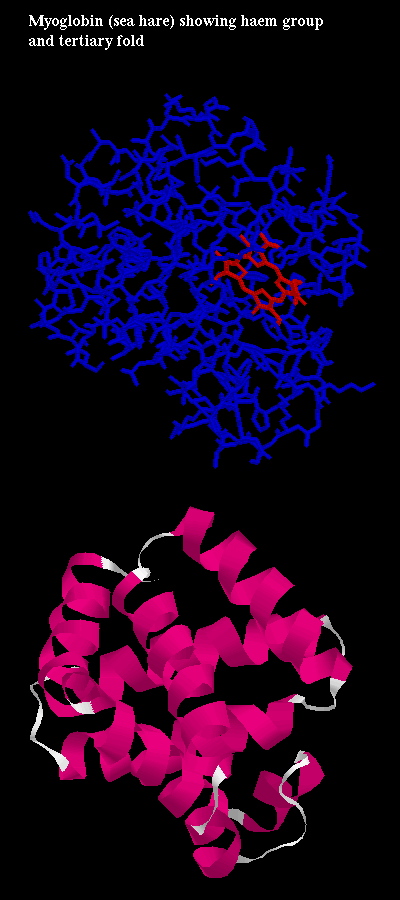 Click here for diagram of
myoglobin, an oxygen-carrying protein which contains a haem porphyrin
(ring) group which has an Fe ion ligand at its centre. It is at this site
that one oxygen molecule is bound. Here is the
PDB structure.
Click here for diagram of
myoglobin, an oxygen-carrying protein which contains a haem porphyrin
(ring) group which has an Fe ion ligand at its centre. It is at this site
that one oxygen molecule is bound. Here is the
PDB structure.
The enzyme carboxypeptidase A is a zinc metallo-protease, which has a single Zn++ ion bound to its single polypeptide chain. This ion is involved in the binding of substrates and inhibitors (see the section on enzymes).Click here for a diagram of bovine pancreatic carboxypeptidase A, showing the zinc ion at the active site. (This picture, and others referred to in this section, were generated by Manuel Peitsch at GLAXO Geneva.)
Insulin exists of a trimer of dimers in a hexagonal arrangement. Each monomer consists of two chains joined by disulphide bridges. There are two forms of insulin, in which either 2 or 4 Zn ions are bound. Click here for the insulin dimer showing zinc ion binding sites.
Several examples of protein-ligand binding are available, produced by Andrew Wallace and Roman Laskowski's LIGPLOT software.
Back to Main PPS Index
J. Walshaw
{kind=link}
{kind=link}