 Analysis of NMR Spectra
Analysis of NMR Spectra
Analysis of NMR Spectra
Analysis of NMR Spectra
The strategies employed for the assignment procedure depend on whether only homonuclear 2D spectra are available (unlabelled proteins), whether 15N heteronuclear spectra are available (15N labelled proteins) or whether triple resonance spectra (15N/13C doubly labelled proteins) are available. But in general the assignment can be divided in two parts: The sequential assignment of the amino acids in the protein sequence and the assignment of the amino acid side chains.
The fist step in sequential assignment is the identification of certain amino acids, with a characteristic pattern of cross signals, i.e. of glycine, alanine, threonine, valine, leucine and isoleucine.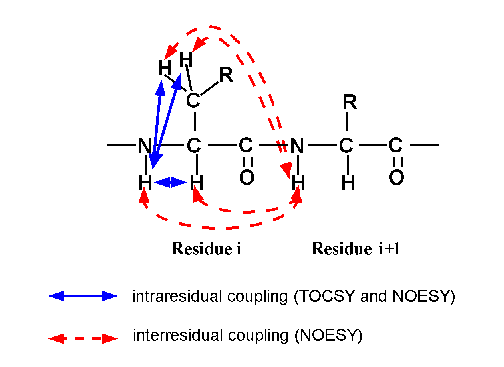
Glycine (left picture) contains two Halpha protons and is therefore readily identified. Valine (right picture), leucine and isoleucine can be recognized by their two methyl groups which give a characteristic row of double signals between 0 and 1.5 ppm. In the same way, alanine and threonine are identified by their single methyl groups.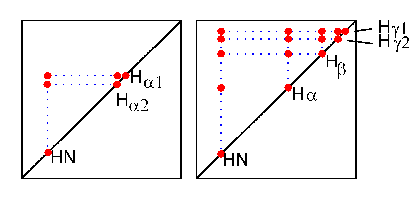
In the second stage of the assignment process, the sequential contacts from the already identified amino acids to the neighboring ones are searched for in the 2D NOESY spectra. The connectivity of a given amino acid in the sequence (i) to its following one (i+1) can be observed in the NOESY because the distance of the amide proton of (i+1) to the Halpha, Hbeta or Hgamma protons of (i) is smaller than 5 A in almost every case (left picture). Therefore, sequential cross signals to Halpha(i), Hbeta(i) etc. are observed at the frequency of HN(i+1) (right picture, dark blue signals). These interresidual cross signals can be distinguished from the intraresidual ones by comparing the 2D NOESY with the 2D TOCSY spectrum. A series of these sequential cross signals between Halpha(i) and HN(i+1) determines the order (i, i+1, i+2,...) of the amino acid spin systems.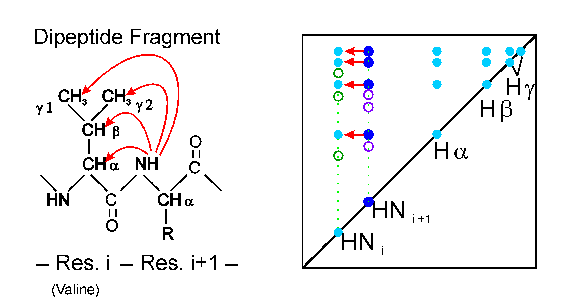
Thus, dipeptides are identified and subsequently prolonged to oligopeptides by the search for further sequential contacts. Some time along the line these oligopeptides can be placed at a unique place in the primary structure by comparison with the amino acid sequence of the protein - they are sequentially assigned.
The chain of sequential connectivites is interrupted by proline residues because these have no amide proton. Therefore, no HN(i)-Halpha(i-1) cross signal can be observed. However, if the proline (i) is in its trans conformation, the sequential HN(i-1)-Hdelta(i) and Halpha(i-1)-Hdelta(i) cross signals can be observed.
Another problem is, that this approach of sequential assignment breaks down for larger proteins because the vast number of signals leads to spectral overlap which hinders the identification of signals.
A HNCA spectrum has three frequency axes: 1H, 15N and 13C. It correlates an amide proton with the Calpha atom of the 'own' and in most cases also with the Calpha of the preceeding amino acid. A 1H/15N projection of a HNCA looks like an HSQC: Each signal represents a single amino acid. At the frequency of each amide proton there are two cross signals in the Calpha dimension: One from the intraresidual and one from the interresidual Calpha atom. Using these cross signals a chain of correlations through the whole amino acid sequence can be established, just like building a chain of dominoes. However, proline residues interrupt that chain due to their missing amide proton.
The assignment is much easier if the intra- and interresidual cross signal can be distinguished. This discrimination can be made using the HN(CO)CA experiment which also correlates amide protons with Calpha atoms. Since the magnetization in the HN(CO)CA is transferred via the CO atom, only the interresidual cross signal can be observed. Therefore, a HNCA and a HN(CO)CA spectrum are laid on one another to discriminate intra- and interresidual Calpha cross signals.
This picture (32 k) illustrates the 'domino pattern' that is obtained during the sequential assignment with triple resonance spectra. It shows superpositions of a HNCA spectrum (blue) and a CBCA(CO)NH spectrum (red) (the latter is an analogue to the HN(CO)CA). The picture containes strips from the two spectra which correspond to a single amino acid. Several of these strips are placed in a row to show the sequential connectivities from each amino acid to the preceeding one (marked with green boxes and arrows).
Usually the sequential assignment by HNCA and HN(CO)CA is ambigous because the Calpha frequencies of totally different amino acids are accidentally degenerate. Thus, the predecessor of a given amino acid often cannot be defined because there are several possibilities. Therefore, correlations via other types of nuclei have to be used for resolution of ambiguities in the HNCA, for example the HNCO and HN(CA)CO experiment. These two spectra correlate the frequency of an amide proton with the frequencies of the intra and interresidual CO atom. They are an independent alternative for checking the sequential connectivities.
Besides, the HNCO is very useful for resolving accidental signal degenerations in the HSQC projection: In proteins every amide proton is covalently bonded to a single CO group. Therefore, only one cross signal per frequency is observed in the HNCO. However, if two cross signals are observed at the frequency of an amide proton there have to be two amino acids with accidentally degenerated amide protons.
Another two pairs of triple resonance experiments which deliver an independent assignment strategy are HBHA(CBCACO)NH and HBHA(CBCA)NH which correlate amide protons via the Ha and Hb frequencies and the closely related experiments CBCA(CO)NH and CBCANH which correlate amide protons via Calpha and Cbeta resonances. The latter have the additional advantage over HNCA/HN(CO)CA, that they have cross signals to the Cbeta atoms of the amino acids providing additional information: In some cases the chemical shift of Calpha and Cbeta atoms can be used for a preliminary identification of the amino acid type.
Of special importance is also the HCACO experiment which correlates the Halpha, Calpha and CO frequencies of an amino acid. If the resonances of different amino acids are degenerate in the HSQC projection, the HCACO is a convenient method to establish the correct combinations of these three frequencies.
The sequential assignment by triple resonance experiments yields only the connectivities of the individual spin sytems with each other but not their amino acid type. The correct amino acid type can be identified with TOCSY-HSQC or HCCH-TOCSY experiments.
Taking it all together, the strategy is as follows:
{kind=link}