![]()
![]()
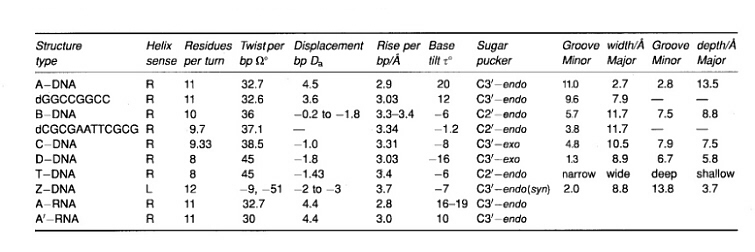
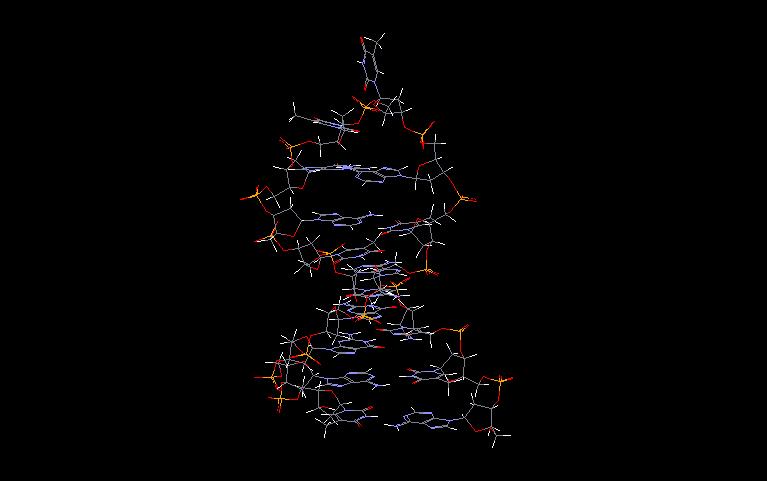
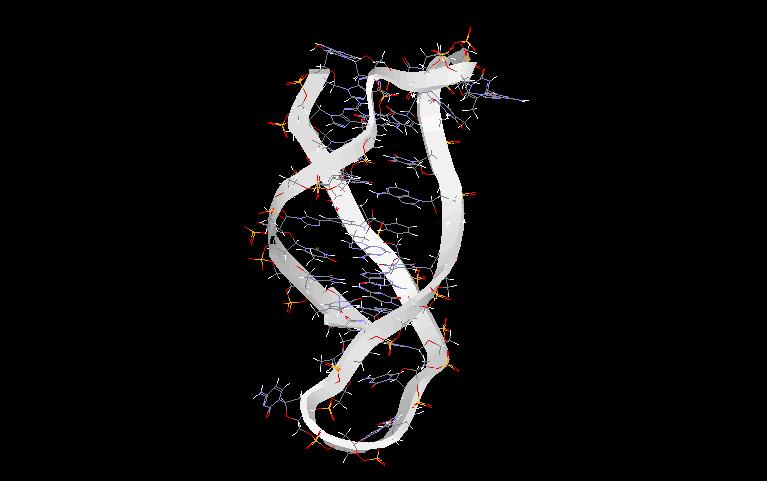
In the first phase of investigating the DNA secondary structure, diffraction studies on heterogeneous DNA identified two distinct conformations of the DNA double helix. At high humidity (and low salt) the dominant structure is called B-DNA (as a jpg file or pdb file) and at low humidity (and high salt) the favoured form is A-DNA (as a jpg file or pdb file). At the present we recognise the presence of a wide variety of right-handed double helical DNA conformations. This structural polymorphism is expressed in different symbols for each structure; known structures of DNA are marked A, A’, B, alpha-B’, beta-B’, C, C’ D, E, T (average helix parameters). In 1979 a quite different structure was discovered namely a left handed helix formed by a synthetic hexamer d(CGCGCG), now known as a Z-DNA (as a jpg file or pdb file). In broad terms we can distinguished three families of DNA structures: A-DNA, B-DNA and Z-DNA. The difference lies primarily in the sugar pucker which is C3’-endo for the A family, C2’-endo for the B family, and C2’-endo for cytosine and C3’-endo for guanine (in conformation syn) in Z-DNA.
Apart from mentioned above structures single strand DNA can form a hairpin (as a jpg file or pdb file) structure, or hybridising to a double helix, a triplex (triple helix) (as a jpg file or pdb file). In some cases two double helices can align to each other generating a new structure called tetraplex (as a jpg file or pdb file).
Some organisms have their DNA organised in circular forms. Prokaryota (and also primitive Eukaryota) apart from a circularised genome have short extrachromosomal DNA molecules (called plasmids) organised in the same way. The connection between a 5’ end a 3’ end leads to the situation where not only the secondary structure of a double stranded DNA but also the superhelical twist usually appearing as the result of enzymatic (topoisomerases I, gyrase, helicase) activity must be taken into account.
As more highly resolved structures have become available, the idea that DNA conformations are restricted to standard structures has whittled away. It is now accepted that there are local, sequence-dependent modulations of structure which are primarily associated with changes in the orientation of bases. Such changes seek to minimize non bonded interactions between adjacent bases and maximise base-stacking. Different base sequences have their own characteristic signature: they influence groove width, helical twist, curvature, mechanical rigidity, and resistance to bending. It seems probable that these features help proteins to read and recognise one base sequence in preference to another.
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}