Guanine nucleotide binding proteins are glycoproteins anchored on the cytoplasmic cell membrane. They are mediators for many cellular processes, including signal transduction, protein transport, growth regulation and polypeptide chain elongation. They are also known as GTP-binding proteins and GTPases. Almost all members of this superfamily of proteins act as molecular switch, which is on when GTP is bound and off when GDP is bound. Binding is specific for the guanine base, with affinity of the order of 1011-1012 M-1 for GDP and GTP, whilst binding affinity for GMP and other nucleotide bases are 6 orders less.
The activity of guanine nucleotide binding proteins is dependent on divalent ions, which is Mg++ in vivo. Mg++ forms octahedral coordinate bonds. When GTP is bound, two neighbouring equatorial dative bonds are connected to 2 oxygen atoms of the beta and gamma phosphates of GTP, whilst the other two equatorial bonds are connected to the hydroxyl oxygen of Ser17 and Thr35, respectively. Both polar dative bonds are linked to water molecules. When GDP is bound, the coordinately bonded oxygen of gamma phosphate is replaced by a water molecule. The hydroxyl oxygen of Thr35 is also broken off and replaced by a water molecule.
Ras proteins are divided into two domains, G domain which is the guanine nucleotide binding region, and the C domain which anchors the protein onto the cytoplasmic side of the cell membrane. Ras proteins belongs to the alpha/beta class of proteins, with the G domain made of a central beta sheet of 6 strands, flanked below by 2 alpha helices and above by 3 alpha helices. These are connected by turns and loops. Loops 1, 2 and 4 are of crucial importance as they surround the active centre at the gamma phosphate group of bound GTP.
![]() 5P21
shows guanosine-5'-(beta, gamma-imido)triphosphate bound to the G domain
of p21Hras. Caged GTP or analogues are used for crystallographic studies
as GTP is hydrolysed during crystallisation. Mg++ is shown as
a green ball. GppNp is shown in ball and stick with CPK colouring. Loops
1, 2 and 4 are shown in violet, magenta and purple colours respectively.
These three loops surround the active centre containing GTP and Mg++.
5P21
shows guanosine-5'-(beta, gamma-imido)triphosphate bound to the G domain
of p21Hras. Caged GTP or analogues are used for crystallographic studies
as GTP is hydrolysed during crystallisation. Mg++ is shown as
a green ball. GppNp is shown in ball and stick with CPK colouring. Loops
1, 2 and 4 are shown in violet, magenta and purple colours respectively.
These three loops surround the active centre containing GTP and Mg++.
GTP-bound p21Hras mediate cellular responses by acting on its effector molecules, like phospholipases C and A2 and adenylyl cyclase. This activity is tightly regulated by hydrolysis of bound GTP into GDP which is subsequently released from the G protein. The GTPase activity may be intrinsic or due to the action of effector molecules. GTPase activity is stimulated by GTPase activiating protein(GAP). The markedly different action of the G proteins on and off state is structurally related to its conformational change between the two states. In the GTP-bound on state, loops 2 and 4 of the G protein are tied into a connected loop conformation by the gamma phosphate of GTP. The connection is strengthened with Thr35 of loop 2 coordinated to Mg++ ion which also coordinates to both the beta and gamma phosphates of GTP.
![]() Loops
2 and 4 connection is effected by Thr35 of loop 2 and Gly60 of loop
4 interacting with GppNp. The amide groups of Thr35 and Gly60 form hydrogen
bonds with 2 oxygen atoms of the gamma phosphate, thus connecting the loops
together. Thr35 is also coordinated to Mg++ through its OH group.
Loops
2 and 4 connection is effected by Thr35 of loop 2 and Gly60 of loop
4 interacting with GppNp. The amide groups of Thr35 and Gly60 form hydrogen
bonds with 2 oxygen atoms of the gamma phosphate, thus connecting the loops
together. Thr35 is also coordinated to Mg++ through its OH group.
The following two diagrams illustrate the close contacts that allow the
hydrogen bons to connect the two loops.
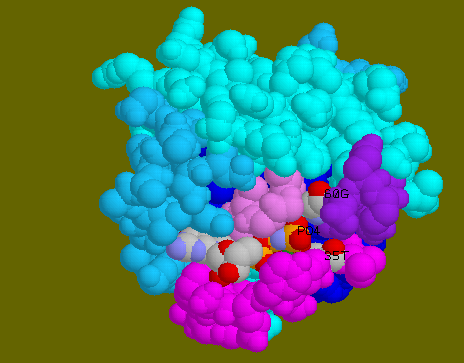
![]() GppNp,
Mg, Thr35 and Gly60 in spacefill model to show the close contact of
gamma PO4 of GppNp with the other three molecules.
GppNp,
Mg, Thr35 and Gly60 in spacefill model to show the close contact of
gamma PO4 of GppNp with the other three molecules.
![]() Whole
complex in spacefill model to confirm the close contact of gamma PO4
of GppNp with the Thr35 and Gly60.
Whole
complex in spacefill model to confirm the close contact of gamma PO4
of GppNp with the Thr35 and Gly60.
When the gamma phosphate is cleaved by nucleophilic attack by water,
loops 2 and 4 are disconnected from each other. Thr35 and loop 2 is also
released from coordination by Mg++ ion. The two loops reorientate
themselves in the GDP-bound state.
The reorientations of loops 2 and 4 are shown in the following diagrams
![]() Loops
in ribbon model to depict the reorientations.
Loops
in ribbon model to depict the reorientations.
![]() GDP,
Thr35 and Gly60 in spacefill model to show that Thr35 and Gly60 are
no longer in close contact with GDP.
GDP,
Thr35 and Gly60 in spacefill model to show that Thr35 and Gly60 are
no longer in close contact with GDP.
![]() Whole
complex in spacefill model emphasising that Thr35 and Gly60 are no
longer in close contact with GDP.
Whole
complex in spacefill model emphasising that Thr35 and Gly60 are no
longer in close contact with GDP.
The effect of this conformational change on the action of guanine nucleotide
binding proteins on the effector molecules is unknown because of the lack
of knowledge on the mechanism of interaction between guanine nucleotide
binding proteins and effectors. This is despite a lot of speculation on
the role of GTPase and GAP as effector molecules.
Much more is known concerning the effect of the conformational change on
the GTPase activity. The gamma phosphate is cleaved by nucleophilic attack
by a water molecule. This nucleophilic attack is assisted when the water
molecule is strongly polarised by forming hydrogen bonds. In the GTP-bound
conformation, Gln61 in loop 4 is in the favourable position for its side
chain carbonyl oxygen to form hydrogen bond with the attacking water molecule.
This polarising effect on the nucleophilic water attacker is further assisted
by hydrogen bonding with the main chain carbonyl oxygen of Thr35 in loop
2, which is favourably oriented in the GTP-bound conformation. After the
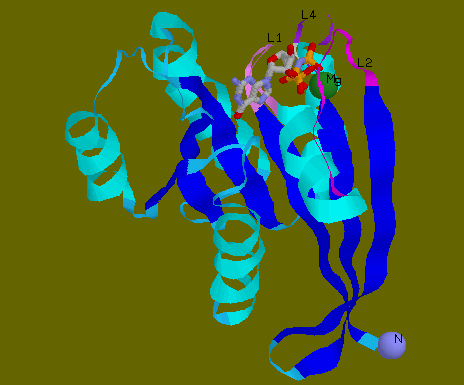{kind=link}
{kind=link}
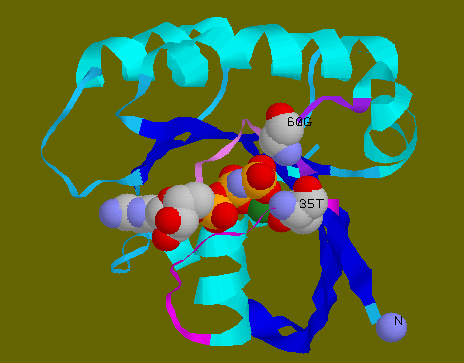{kind=link}
{kind=link}
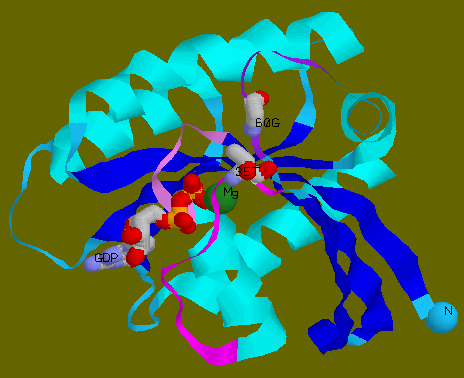{kind=link}
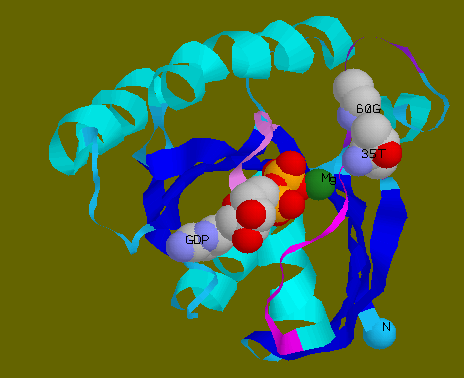{kind=link}
{kind=link}