The major histocompatibility complex (MHC) was originally discovered as transplantation antigens that predominantly determine the compatibility of tissues between different individuals. The complex is comprised of three classes of genes. Classes I and II genes encode antigens expressed on cell surface, whilst class III genes encode several components of the complement system. Classes I and II antigens are glycoproteins that present peptides to T lymphocytes.
Human and murine class I molecules are heterodimers, consisting of a heavy alpha chain (45kD) and a light chain, beta-2-globulin (12kD). Alpha chain can be divided into three extracellular domains, alpha1, alpha2 and alpha3, in addition to the transmembranous and cytoplasmic domains. The alpha3 domain is highly conserved, as is beta-2-microgloobulin. Both alpha3 domain and beta-2-microglobulin are homologous to the CH3 domain of human immunoglobulin.
Class II molecules are heterodimeric glycoproteins, alpha chain (34kD) and beta chain (29kD). Each chain has 2 extracellular domains, togethere with the transmembranous and cytoplasmic domains. The membrane-proximal alpha2 and beta2 domains are homologous to immunoglobulin CH domain.
There are 3 class I loci (B,C,A) in the short arm of human chromosome 6, and 4 loci (K, D(L), Qa, Tla) in murine chromosome 17. These loci are highly polymorphic. The variable residues are clustered in 7 subsequences, 3 in alpha1 domain and 4 in alpha2 domain. There are 3 major human class II loci (HLA-DR, HLA-DO, HLA-DP) and 2 murine loci (H-2I-A, H-2I-E). All class II beta chains are polymorphic. Human HLA-DQ alpha chain is also polymorphic.
Class I molecules present peptides to CD8-CD4+ cytotoxic T lymphocytes, whilst class II molecules present peptides to CD4+CD8- T helper lymphocytes. The composite peptide-MHC macromolecule is recognised by T-cell receptor (TCR) on the surface of T lymphocytes. The problem for the MHC molecules is to present a vast variety of peptides for specific recognition by TCR, with only a limited isotypes of each class in an individual. In terms of molecular structure, many peptides are to be pasted onto a single MHC dimeric protein. The consequence of these peptides pasting is the creation of different conformations of composite peptide-MHC macromolecules. In purely structural terms, one can view the peptide-MHC composite as a trimeric protein. Indeed, in the case of class I molecule, bare dimers are unstable. Class I molecules are stabilised by peptide binding and only the trimeric forms are expressed on cell surface. Classes I and II molecules are versatile in making a large variety of specifically distinct conformations by pasting different peptides onto themselves.
The molecular structure of unbound class I molecule is revealed by the crystallographic structure of the extracellular domain of HLA-A2 (cleaved by papain) at 2.6 Angstrom. There is a peptide-binding groove formed by 2 helices lying parallel on a floor formed by 8 beta strands in alpha1 and alpha2 domains.
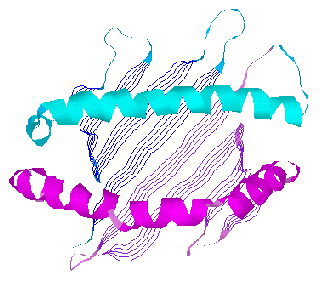
![]() Ribbon model of HLA-A2.1 binding groove viewed from the top (alpha3
domain and beta-2-microglobulin not shown): alpha1 domain helix(cyan),
sheet(blue), turns(dark cyan); alpha2 domain helix(magenta), sheet(purple),
turns(violet)
Ribbon model of HLA-A2.1 binding groove viewed from the top (alpha3
domain and beta-2-microglobulin not shown): alpha1 domain helix(cyan),
sheet(blue), turns(dark cyan); alpha2 domain helix(magenta), sheet(purple),
turns(violet)
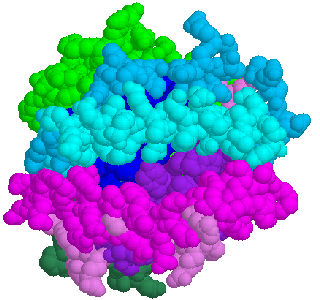
![]() Spacefill model of HLA-A2.1 binding groove viewed from the top: alpha3
domain(greenblue); beta-2-microglobulin(green<. Part of the floor of
the beta sheet can be seen through the groove(blue and purple)
Spacefill model of HLA-A2.1 binding groove viewed from the top: alpha3
domain(greenblue); beta-2-microglobulin(green<. Part of the floor of
the beta sheet can be seen through the groove(blue and purple)
![]() Ribbon model of HLA-A2.1 viewed from the side: Cysteines in disulphide
bonds are shown in yellow
Ribbon model of HLA-A2.1 viewed from the side: Cysteines in disulphide
bonds are shown in yellow
![]() Spacefill model of HLA-A2.1 viewed from the side: The membrane-proximal
portion consists of alpha3 domain and beta-2-globulin on the left of the
diagram. Class I molecule is anchored onto the cell membrane through its
alpha chain. The model shown is truncated at residue 270, where the transmembranous
domain is connected.
Spacefill model of HLA-A2.1 viewed from the side: The membrane-proximal
portion consists of alpha3 domain and beta-2-globulin on the left of the
diagram. Class I molecule is anchored onto the cell membrane through its
alpha chain. The model shown is truncated at residue 270, where the transmembranous
domain is connected.
Class I molecules usually bind nonapeptides, and less frequently octapeptides
or decapeptides. Nonapeptides bind in an extended conformation with a kink
near P4. Octapeptides bind with less acute kink, whilst decapeptides and
longer peptides bind with more pronounced kinks or zigzags in the extended
conformation.
The peptides are anchored onto "pockets" in the peptide-binding
groove of MHC dimers only at a few positions of the peptides. These positions
are called "anchors", and are usually at P2 or P3 and Psigma.
Other side chains of the bound peptide also make contact with the groove,
but not apparently constrained by any specific pocket.
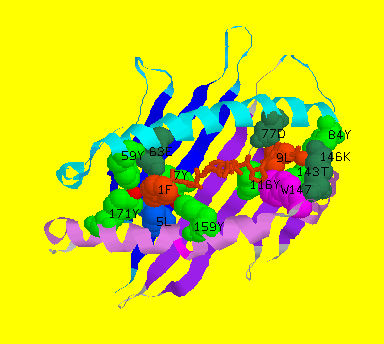
![]() Diagram of Sendai virus nucleoprotein nonapeptide-bound HLA-2kb, showing
pockets A and F. Pocket A accomodates the N-terminus of the peptide with
three highly conserved tyrosines at residues 7, 159 and 171 that form hydrogen
bonds with the amide group and carbonyl oxygen of the first peptide residue.
Pocket F holds the C-terminus where the carboxyl group of the last residue
and the carbonyl oxygen of the penultimate residue make extensive hydrogen
bonding with highly conserved side chains of Tyr84, Thr143, Lys146 and
Trp147 and with the moderately conserved side chain of Asp77.
Diagram of Sendai virus nucleoprotein nonapeptide-bound HLA-2kb, showing
pockets A and F. Pocket A accomodates the N-terminus of the peptide with
three highly conserved tyrosines at residues 7, 159 and 171 that form hydrogen
bonds with the amide group and carbonyl oxygen of the first peptide residue.
Pocket F holds the C-terminus where the carboxyl group of the last residue
and the carbonyl oxygen of the penultimate residue make extensive hydrogen
bonding with highly conserved side chains of Tyr84, Thr143, Lys146 and
Trp147 and with the moderately conserved side chain of Asp77.
![]() Diagram of Sendai virus nucleoprotein nonapeptide-bound HLA-2kb, showing
pocket C. Pocket C is deep in murine H-2k, compared to the corresponding
human HLA-A molecule.
Diagram of Sendai virus nucleoprotein nonapeptide-bound HLA-2kb, showing
pocket C. Pocket C is deep in murine H-2k, compared to the corresponding
human HLA-A molecule.
Each class I dimeric molecule presents different conformation to the
TCR, when different peptides are bound on it. It is shown by crystallographic
structure of at least nine human peptide-bound HLA-A isotype and three
murine peptide-bound H2k isotype that conformational changes occur in the
neighbourhood of the peptide-class I interface. The bound peptides are
in an extended secondary structure showing distinct conformational changes
with different peptides. The class I dimer does not show significant conformational
changes, except for some minor changes in several residues interacting
with the peptides. These data appear to indicate that bound peptides do
not induce allosteric changes in conformation.
The central portion of the bound peptide shows most striking changes in
conformation. This is also the region exposed on the trimeric complex,
and hence readily availabel for TCR recognition. Depending on the sidechains
present in the peptide, the main chain zigzag within the binding groove
in different course. The zigzag in turns affects the orientaion of the
sidechains of the peptide. This differential sidechain orientation is most
pronounced in the central portion, as the extreme residues are anchored
at both ends of the groove.
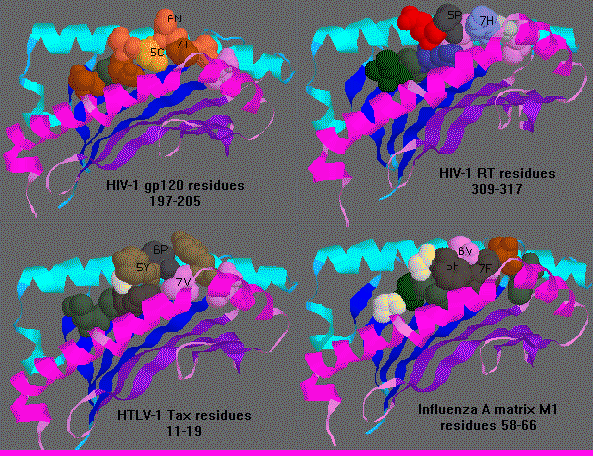
The following diagram shows the variation in the twist and turn and side
chain orientations of residues 5-7 (P5,P6,P7) in HLA-A0201 bound by 4 different
nonapeptides.
![]() Diagram of 4 nonapeptide-bound HLA-A0201. P5 points up in HTLV-1 tax
and HIV-1 RT, but points down in HIV-1 gp120 and points sideway toward
the alpha2 alpha helix in Influenza A matrix. P6 points up in HIV-1 gp120
and Influenza A matrix, down in HIV-1 RT and sideway toward the alpha1
alpha helix in HTLV-1 tax. P7 points up in HIV-1 RT, down in Influenza
A matrix, but sideway toward to alpha2 alpha helix in HIV-1 gp120 and HTLV-1
tax.
Diagram of 4 nonapeptide-bound HLA-A0201. P5 points up in HTLV-1 tax
and HIV-1 RT, but points down in HIV-1 gp120 and points sideway toward
the alpha2 alpha helix in Influenza A matrix. P6 points up in HIV-1 gp120
and Influenza A matrix, down in HIV-1 RT and sideway toward the alpha1
alpha helix in HTLV-1 tax. P7 points up in HIV-1 RT, down in Influenza
A matrix, but sideway toward to alpha2 alpha helix in HIV-1 gp120 and HTLV-1
tax.
Class I molecules are unique in exhibiting conformational varieties by pasting onto its groove a selection of peptides.
{kind=link}
{kind=link}
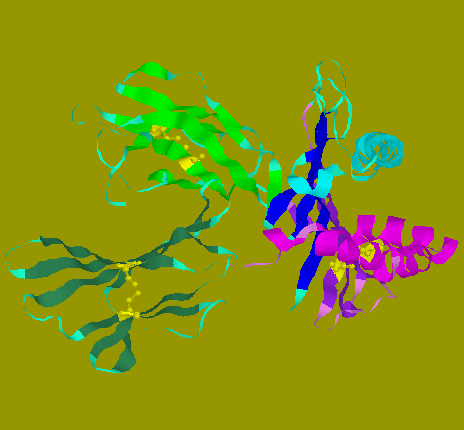{kind=link}
{kind=link}
{kind=link}
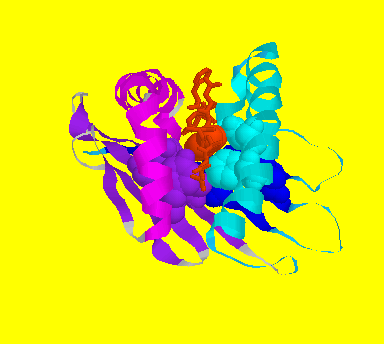{kind=link}
{kind=link}