Alcohol dehydrogenases oxidises alcohol into aldehydes orketones. The reaction requires conenzyme NAD+ as hydrogen acceptor and has a broad specificity for alcohol substrates. They belong to a family of dehydrogenases that have a nucleotide-binding domain. The strategy of all such NAD-dependent dehydrogenases is to orient the coenzyme and substrate on the enzyme surface, such that the C4 atom on the nicotinamide is directed at the reactive carbon of the substrate. The NAD-binding domains of all these dehydrogenases are highly homologous, but they have markedly different catalytic domains.
Alcohol dehydrogenase is a homodimer. Each monomer has 374 residues with molecular weight of 74000 dalton. There are two domains. The NAD+-binding domain (residues 176-318) consists of a central beta-sheet of 6 strands flanked by alpha helices. NAD+ binds to the C-terminus of the beta-sheet. The catalytic domain (residues 1-175, 319-374) also has a alpha/beta structure. The inter-domain interface forms a cleft which contains the active catalytic site. The interface is formed by two helices, one from each domain crossing over each other. There are two Zn++ cations per monomer, one at the catalytic site being mandatory for catalysis. The alcohol substrate binds inside the cleft where the Zn++ cation is, whilst the nicotinamide ring of the NAD finds its way pointing into the cleft. The dimer forms with the two NAD-binding domains packing together such that their 2 central beta sheets combine to form a 12-stranded beta sheet. The catalytic domains are situated at opposite ends.
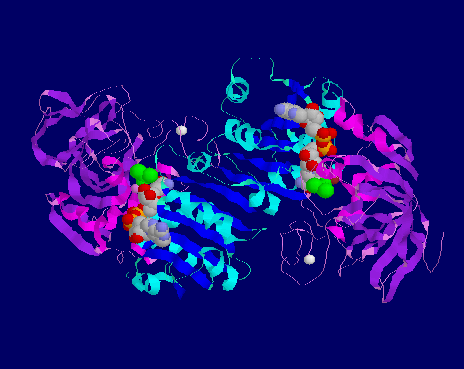
![]() 6ADH-
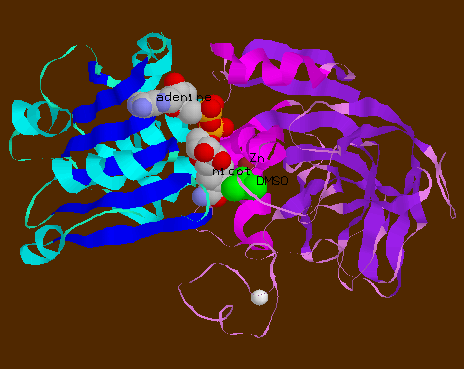
Ribbon view of horse liver alcohol dehydrogenase monomer The NAD+-binding
domain is shown with helix in cyan and sheet in blue. The catalytic domain
has helix in magenta and sheet in purple. The substrate is dimethylsulphoside
(DMSO) in green. The active Zn++ ion is in brown, whilst the
other one is in white. NAD+ is in CPK colouring, with its adenine
ring bound to the C-terminus of the beta sheet of the NAD+-binding
domain. Its nicotinamide ring is brought into close proximity with the
substrate and Zn++ ion.
6ADH-
Ribbon view of horse liver alcohol dehydrogenase monomer The NAD+-binding
domain is shown with helix in cyan and sheet in blue. The catalytic domain
has helix in magenta and sheet in purple. The substrate is dimethylsulphoside
(DMSO) in green. The active Zn++ ion is in brown, whilst the
other one is in white. NAD+ is in CPK colouring, with its adenine
ring bound to the C-terminus of the beta sheet of the NAD+-binding
domain. Its nicotinamide ring is brought into close proximity with the
substrate and Zn++ ion.
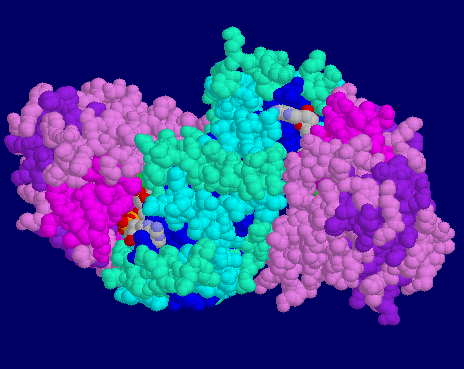
![]() 6ADH-
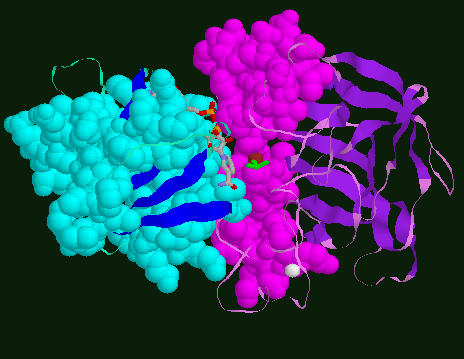
Crossing helices shown in van der Waals spaceballs to highlight the
crossing of helices between the two domains. NAD+ and DMSO are
shown in stick model.
6ADH-
Crossing helices shown in van der Waals spaceballs to highlight the
crossing of helices between the two domains. NAD+ and DMSO are
shown in stick model.
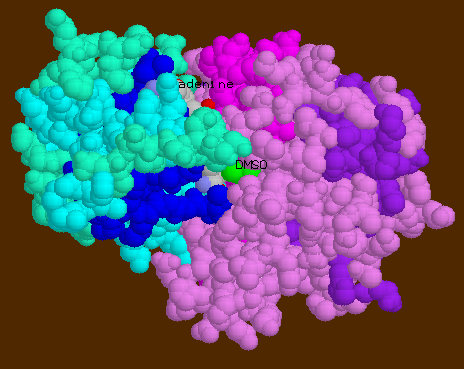
![]() 6ADH-_DMSO
can be seen buried within the cleft in spacefill model.
6ADH-_DMSO
can be seen buried within the cleft in spacefill model.
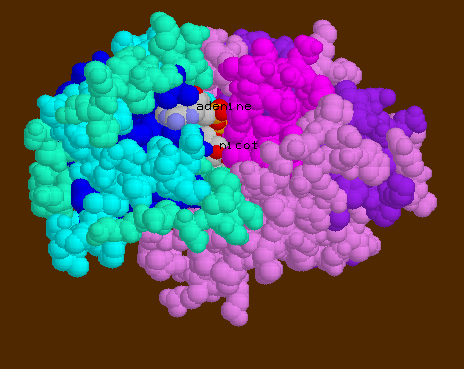
![]() 6ADH-
NAD is seen with its nicotinamide ring buried inside the cleft where
it interacts with the substrate and Zn++ ion.
6ADH-
NAD is seen with its nicotinamide ring buried inside the cleft where
it interacts with the substrate and Zn++ ion.
When the apo-enzyme is bound by its substrate and coenzyme, there is a conformational change. This consists of a rotation of about 7.5 around a hinge axis passing through the contact point of the alpha helices connecting the two domains. This domain motion is classified as a shear motion according to Chothia and Lesk's classification of domain motions. This results in a change in the shape of the cleft to accommodate the substrate. The result is closure of the cleft with the substrate and nicotinamide ring surrounding the Zn++ ion. The active Zn++ ion is in tetrahedral coordination. In the apo-enzyme one of its four ligands is a water molecule. In the holo-enzyme the water molecule is replaced by the hydroxyl oxygen of the alcohol substrate, as a result of solvent exclusion upon cleft closure.
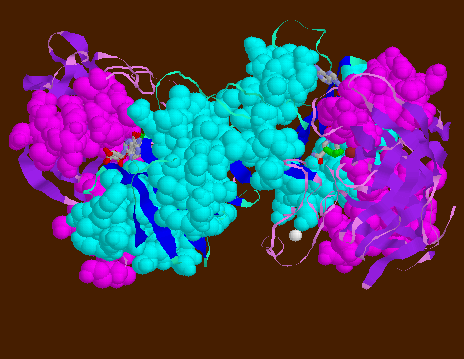
![]() 6ADH-
Dimer helical shear is highlighted in this view with the helices in
spacefill model. The two catalytic domains can be seen with their helices
(magenta) wrapping over the central combined alpha/beta structure of 12-stranded
beta-sheet and flanking alpha-helices (cyan). With the crossed helices,
the catalytic domains can glide over the NAD+-binding domains.
6ADH-
Dimer helical shear is highlighted in this view with the helices in
spacefill model. The two catalytic domains can be seen with their helices
(magenta) wrapping over the central combined alpha/beta structure of 12-stranded
beta-sheet and flanking alpha-helices (cyan). With the crossed helices,
the catalytic domains can glide over the NAD+-binding domains.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}