![]() Index to Course Material
Index to Course Material
![]() Index to Section 9
Index to Section 9
![]() Index to Super Secondary Structure
Index to Super Secondary Structure
![]() Previous Page
Previous Page
Antiparallel beta-strands can be linked by short lengths of polypeptide forming beta-hairpin structures. In contrast, parallel beta-strands are connected by longer regions of chain which cross the beta-sheet and frequently contain alpha-helical segments. This motif is called the beta-alpha-beta motif and is found in most proteins that have a parallel beta-sheet. The loop regions linking the strands to the helical segments can vary greatly in length. The helix axis is roughly parallel with the beta-strands and all three elements of secondary structure interact forming a hydrophobic core. In certain proteins the loop linking the carboxy terminal end of the first beta-strand to the amino terminal end of the helix is involved in binding of ligands or substrates. The beta-alpha-beta motif almost always has a right-handed fold as demonstrated in the figure.
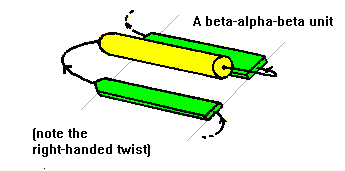
The subsequent sections of this course on protein folds and domains will demonstrate how the beta-alpha-beta motif is an important and widespread element of supersecondary structure.
Introduction to Protein Structure, C.Branden and J.Tooze, Garland, New York (1991).
A novel super-secondary structure of proteins and the relation between the structure and amino acid sequence, A.F.Efimov, FEBS Lett. 166, 33-38 (1984).
Structure of alpha-alpha-hairpins with short connections, A.V.Efimov, Protein Engin.4, 245-250 (1991).
Structure of beta-beta-hairpins and beta-beta-corners, A.V.Efimov, FEBS Lett., 284, 288-292 (1991).
Structure and evolution of calcium modulated proteins, R.H.Kretsinger, CRC Crit.Rev.Biochem. 8, 119-174 (1980).
Comparison of the structures of cro and lambda repressor proteins in bacteriophage lambda, D.H.Ohlendorf, W.F.Anderson, M.Lewis, C.O.Pabo and B.W.Matthews, J.Molec.Biol. 169 757-769 (1983).
Conformation of beta-hairpins in protein structures, B.L.Sibanda, T.L.Blundell and J.M.Thornton, J.Molec.Biol. 206, 759-777 (1989).
Beta-hairpin families in globular proteins, B.L.Sibanda and
J.M.Thornton, Nature 316, 170-174 (1985).
j.cooper 26/1/95
Last updated 31st May '96