
3.1 Helices
A 12 residue alpha helix will contain only 8 hydrogen bonds, despite the 12 backbone NH (donors) and 12 backbone CO (acceptors). The N- and C-terminal ends of an isolated helix contain four NH donors and four CO acceptors each, respectively due to edge effects (Figure 2 ). It has been shown that hydrogen bond partners for these otherwise unsatisfied first four amide protons and last four carbonyl oxygens are often provided by polar groups that flank the helix termini (Presta & Rose, 1988; Richardson & Richardson, 1988). Furthermore, these residue positions were found to contain differentiated amino acid preferences. Helix boundary residues (the first and last helical residues) are called Ncap and Ccap at the N- and C-terminus, respectively (Richardson & Richardson, 1988). The proposed nomenclature uses the following definition:
...-N''-N'-Ncap-N1-N2-N3-............-C3-C2-C1-Ccap-C'-C''-...
where N1 through C1 are residues which contain alpha-helical phi, psi angles and the primed residues belong to the residues outside the helix on either end. The eight terminal residues lacking hydrogen bond partners are then Ncap-N3 and Ccap-C3. In their survey of 13 proteins containing 54 alpha helices, 48% of the Ncap-N1-N2-N3- residues and 35% of the C3-C2-C1-Ccap- residues were satisfied by local side chain or backbone hydrogen bonds (as nearly all potential hydrogen bonding groups in a protein are satisfied, the remaining are satisfied by amino acids brought into position by the folding of the protein or by solvent molecules). Strong amino acid preferences are shown in the table below:
Typical Ncap interactions involve a hydrogen bond from the side chain hydroxyl oxygen of Ser and Thr or side chain carbonyl oxygen of Asn with the backbone amide nitrogen of N3. A special form of Ncap utilizing a reciprocal hydrogen bonding motif is called the "capping box" and is shown in Figure 3 . Here the hydroxyl oxygen forms a standard Ncap hydrogen bond with the amide of N3 and the side chain carbonyl oxygen of N3 forms a hydrogen bond with the amide of Ncap thus satisfying two of the four non hydrogen-bonded helix N-terminal amides.
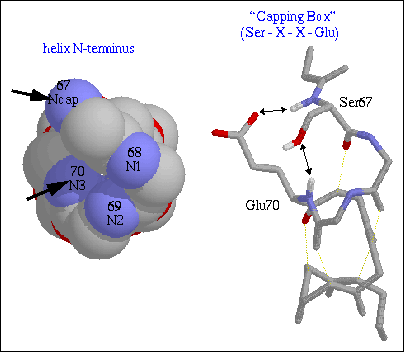
Figure 3. A typical "capping box" from cytochrome 351. On the left is a CPK representation (without sidechains) looking down the helical axis from the N-terminus showing the four non hydrogen-bonded amide nitrogens. On the right, the helix is again shown this time using stick representation following a 90 degree rotation. The four non hydrogen-bonded amide nitrogens are colored blue and the side chains of Ncap and N3 are also shown. The "capping box" hydrogen bonds are shown as black arrows, whereas the helical hydrogen bonds are shown as yellow dashed lines.
Approximately one-third of all alpha helices contain a Gly-based capping motif at the C-terminus. Helices terminating in Gly can be classified into to primary motifs based on their hydrogen bonding patterns. One motif (Schellman motif) contains two "capping" hydrogen bonds, one connecting the backbone amide of C'' to the backbone carbonyl oxygen of C3 and he other connecting the backbone amide of C' to the backbone carbonyl oxygen of C2 (Figure 4 ). The other motif (alpha-L) contains one hydrogen bond connecting the backbone amide of C' to the backbone carbonyl oxygen of C3. In each of these motifs, the C' residue must adopt a left-handed helical conformation which is why Gly is favored although residues with beta carbons are found. See also "Paperclips" .
download cytochrome c351
3.1.4.2 amphiphilicity
Of the forces that drive a polypeptide chain into a folded form in water, solvent interactions (e.g., the hydrophobic effect) are thought to be among the most important (e.g., see Dill, 1990). On a crude level, the folded protein can be thought of as a type of single chain micelle achieving solubility (and stability) by surrounding a largely hydrophobic core with largely hydrophilic exterior.
Figure 5. The "oil-drop" or "micelle" model of globular protein structural organization. On a crude level, a globular protein can be viewed as an oil drop or micelle in which hydrophobic sidechains (balls on the model unfolded protein on the right) are buried away from solvent contact and hydrophilic sidechains (triangles and squares) form the solvent interface.
Helices are very often located parallel to the surface of globular proteins making them prime candidates for serving the function of a barrier between inside and out. As such they should necessarily be amphiphilic (or as some would say, amphipathic) with a hydrophobic face (to satisfy the core) and a hydrophilic face (to interact favorably with the solvent). This is exactly what is found in many helices contained in globular proteins. It was first recognized in myoglobin and since then, numerous methods (see also Figure 32 )for their identification have been proposed (Perutz, 1956; Schiffer & Edmundson, 1967; Finer-Moore & Stroud, 1984; Eisenberg, et al., 1982) and possible functions of these amphiphilic structures have been proposed and experimentally tested (Kaiser & Kézdy, 1984; DeGrado, 1988). An example of a classical amphiphilic alpha helix is found in the four-helix bundle protein cytochrome B562 (Figure 6 ). See also "stereochemical" methods of secondary structure prediction.
The organizing potential of peptides whose amino acid sequence is capable of amphiphilic segregation of side chains when in a helical conformation is quite remarkable. Peptides containing the helical segments of these potential amphiphilic structures will spontaneously adopt the amphiphilic conformation if provided with an amphiphilic environment (e.g. micelle binding, partitioning at the air-water interface). These peptides have been shown to undergo a highly cooperative monomer-oligomer equilibrium at micro-molar concentrations in aqueous solution. At low concentration they are typical random conformers which become helical as the concentration is increased and oligomers (usually tetramers) are formed. Thus amphiphilic alpha helices are a natural starting point for protein design projects (e.g., DeGrado, 1988).
3.1.4.3 helix geometry
In Figure 2 , geometrically pure helices are displayed for clarity, however most (up to 85%) "real" helices in globular proteins deviate from this ideal (Barlow & Thornton, 1988). We have already seen that the phi and psi angles taken from helices identified in three-dimensional structures are slightly different from the ideal values. Helix curvature is another common deviation found in globular proteins. A majority of regular helices contain some curvature of the helical axis (~60 Angstrom radius). Very often this curvature is centered on the hydrophobic side of the helix resulting in slightly different phi and psi angles for the hydrophobic and hydrophilic faces. This curvature is not expected to be energetically expensive and has been estimated to involve < 2 kcal for one mole of a 5-turn helix (Barlow & Thornton, 1988).
Another common distortion to naturally occurring alpha helices is a kink in the helical axis (~25 degrees from linear) caused by the presence of a proline in the interior of the helix. Proline kinked helices are almost exclusively long helices (>4 turns) which might be a requirement to overcome the effect of the kink. Somewhat surprisingly, only one hydrogen bond (amide of the following residue, i+1, to the carbonyl oxygen of i-3) was found to be broken due to the kink in the helix.
The effect of the pyrrolidine ring on the conformational preferences of proline are commonly misunderstood. Since the dihedral angle psi in proline is relatively fixed at about -60 degrees it differs from helical psi angles by about 20 degrees. The ring also has a significant effect on phi resulting in two minima (phi = -55 and phi = 145). Thus proline is largely constrained to conformers, one of which is approximately helical (-55, -60). However proline does exert an effect on the proceeding residue due to the steric interference of the delta carbon (attached to imide nitrogen) with the beta carbon of the proceeding residue. The result is that the dihedral angle, psi, of non-glycine residues proceeding prolines is restricted to positive values (+30 < psi < +180). This (and the proline phi and psi restriction) leads to the kink in proline-containing helices. If the proceeding residue is a glycine, there is no such effect. Thus, the too commonly quoted property of proline as a "helix breaker" is not entirely true. Proline is often found in the first turn of helices (Figure II ) as it's "helix-like" backbone dihedral angles can get the helix started and it is its own capping residue (imide).
3.1.4.4 helix dipole
In a helix, all of the peptide units point in the same direction (roughly parallel to the helical axis) and therefore should show a cumulative effect resulting in a helix macrodipole (Hol et al., 1978). The effect of this helix macrodipole is the presence of approximately +0.5 unit charge at the N-terminus and -0.5 unit charge at the C-terminus. This would explain the observed preferences (Table II ) of negatively charged amino acids at the N-terminus and positively charged residues at the C-terminus of helices. In addition, phosphate moieties are found to bind frequently at the N-termini of helices and a number of enzymes are found to have their active site close to the N-terminal of an alpha helix.
Figure 7. Part of a polypeptide chain (extended conformation) showing the direction of a single amide dipole moment (left) and a cartoon of an alpha helix illustrating the (approximate) individual amide dipoles and the overall helical macrodipole. The partial charges on the amide hydrogen and carbonyl oxygen are shown in units of the elementary charge contributing to an overall dipole moment of 3.46 Debye units (equivalent to a 0.5 unit charge separated by 1.5 Angstroms).
Generated with CERN WebMaker

![]()
![]()
![]()
No Title - 31 MAY 96
written by Kurt D. Berndt