3.0 Types of Secondary Structure
Turns were first identified by Venkatachalam (1968) who found three types each containing a hydrogen bond between the carbonyl oxygen of residue i and the amide nitrogen of i+3. These three types of turns are designated I, II, and III. Type III is simply a single turn of 3.10 helix (3.1.2). Each of these three types of turns has a backbone mirror-image conformation that is also found in some proteins designated I', II', and III' even though these conformations are disfavored due to steric hindrance.
Definitions of turns can be grouped into two classes (see Rose et al., 1985 for a comprehensive review): reference, based on stereochemical templates and requirement for a hydrogen bond (e.g., Venkatachalam's definition) and working: that used by anyone who has looked for turns in globular protein structures, based on any of a number of "relaxed" criteria. To illustrate the problem, Rose et al. (1985) tabulated 20 "working definitions of turns!
Many have speculated on the role of this type of secondary structure in globular proteins. Perhaps the two extremes can be classified as passivists and activists. The passivists view turns as a weak link in the polypeptide chain, allowing the other secondary structures (helix and sheet) to determine the conformational outcome. The activists, (encouraged by the recent experimental finding of "turn-like" structures in short peptides in aqueous solution (Dyson et al., 1988)) view turns as structure nucleating segments, formed early in the folding process. Like all extremes, neither is accurate and evidence can be given to support each view.
Type I turns occur most frequently (2-3 times more frequently than type II). The mirror-image types I' and II' are rare but type I' appears to be preferred in beta-hairpins (Creighton, 1993).
There are position dependent amino acid preferences for residues in turn conformations. Type I can tolerate all residues in positions i to i+3 with the exception of Pro at position i+2. Proline is favored at position i+1 and Gly is favored at i+3 in type I and II turns. The polar sidechains of Asn, Asp, Ser, and Cys often populate position i where they can hydrogen bond to the backbone NH of residue i+2. Ideally, type I' turns have Gly at positions i+1 and i+2 and type II' turns have Gly at position i+1 as the presence of a CB atom would cause a steric clash with the peptide carbonyl oxygen.
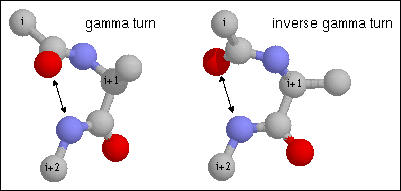
Figure 14. Gamma turn. Note the hydrogen bond between CO of residue i and NH of residue i+2. The dihedral angles of residue i+1 are (70, -60) and (-70, 60) for phi and psi of the classical and inverse gamma turns.
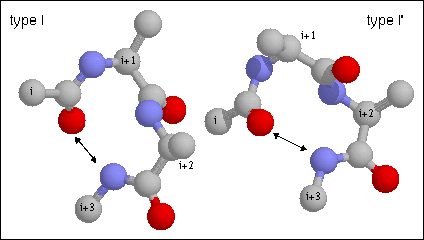
Figure 15. Type I turn. Note the hydrogen bond between CO of residue i and NH of residue i+3. The backbone dihedral angles of residue are (-60, -30) and (-90, 0) of residues i+1 and i+2, respectively of the type I turn. Proline is often found in position i+1 in type I turns as its phi angle is restricted to -60 and its imide nitrogen does not require a hydrogen bond. Glycine is favored in this position in the type II' as it requires a positive (left-handed) phi value.

Figure 16. Type II turn. Note the hydrogen bond between CO of residue i and NH of residue i+3. The backbone dihedral angles of residue are (-60, 120) and (80, 0) of residues i+1 and i+2, respectively of the type II turn. Proline is often found in position i+1 in type I turns as its phi angle is restricted to -60 and its imide nitrogen does not require a hydrogen bond. Glycine is favored in this position in the type II' as it requires a positive (left-handed) phi value.
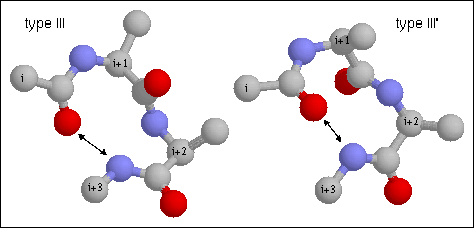
Figure 17. Type III turn. Note the hydrogen bond between CO of residue i and NH of residue i+3. This is a single turn of right-handed (III) and left-handed (III') 3.10 helix, respectively. The backbone dihedral angles of residue are (-60, -30) and (-60, -30) of residues i+1 and i+2, respectively of the classical type III turn.
Generated with CERN WebMaker