 Overview of Enzymes
Overview of Enzymes
 Index to Course Material
Index to Course Material
 Index to Section 12
Enzymes Index;
Index to Section 12
Enzymes Index;
Enzymes are biological catalysts- more than 1500 are known, which catalyze
many different
biochemical pathways. The great majority are confined to the cytosol
(reducing environment).
Consider a chemical reaction in which substrate S is converted to
product P, by an enzyme E.
Enzymes associate with their substrates to form an enzyme-substrate complex
(ES).
In the simplest model (the Michaelis-Menten model):
- E and S associate with a rate constant k1,
- ES dissociates back to E and S with a constant k2
- ES reacts to form E and P with a constant k3 (the turnover number).
The least stable species on the reaction pathway is
called the transition state. Catalysis involves lowering the energy
of this transition state; as a result the rate of the reaction may be increased
over a million-fold compared to the uncatalysed process.
Enzymes bind their substrates more tightly in the transition state
than in their normal states, ie the transition state ES is stabilized.
For a detailed treatment of enzyme kinetics, refer to Fersht (1984).
Biochemical Pathways
A very large number of types of chemical reactions are involved in an
organism's metabolic processes, catalyzed by many different enzymes. An
example of a biochemical pathway is glycolysis,
in which glucose is converted to pyruvate, producing ATP.
Classification of enzymes by function
There are 6 major classes of enzymes:
- Oxidoreductases which are involved in oxidation, reduction, and
electron or proton transfer reactions;
- Transferases, catalysing reactions in which groups are transferred;
- Hydrolases which cleave various covalent bonds by hydrolysis;
- Lyases catalyse reactions forming or breaking double bonds;
- Isomerases catalyse isomerisation reactions;
- Ligases join substituents together covalently.
Enzyme Databases
A more detailed classification is available in
You will notice that the classification is quoted as an E.C. number in the
COMPND record of the PDB file of an enzyme structure.
Active sites
Active sites are the clefts or crevices to which the substrate binds.
In many cases water is excluded from these clefts, which may have a largely
hydrophobic surface, although there will usually be a number of polar
side chains which effect the binding and catalysis of the substrate. In some cases
water molecules are reactants. Many enzymes are highly specific in terms of the
substrate(s) which they bind, and this is determined by the arrangement of
the atoms in the cleft. The binding of a substrate to its active site is often
likened to a key fitting a lock. However the substrate may actually induce a
change in conformation of the active site that allows its accommodation; thus
an induced fit. Substrates are bound to enzymes generally
relatively weakly, often by hydrogen bonds:
ES complexes have equilibrium constants ranging from one hundredth to one
hundred millionth M, which corresponds to interaction free energies of -3 to
-12 kcal/mol (c.f. covalent bonds: -50 to -110 kcal/mol).
Only a small proportion of the enzyme's volume constitutes the active site, and
therefore only a small fraction of the residues of the polypeptide chain are in
contact with the substrate. The residues which constitute
the active site are often not close to each other in the primary sequence, but
the tertiary fold brings them close together in three-dimensional space.
Active sites often involve residues on connecting loops between helices and
sheets, rather than those which are part of these regular secondary structures.
In many instances, active sites occur at the junction between two domains making
tertiary contacts. This is the case in the trypsin family of serine proteases,
examined in more detail later in this section.
Diagram (28Kb GIF) of lysozyme showing active site cleft.
Inhibitors and effectors
The activity of an enzyme can be controlled by the regulation of its
concentration. This may be achieved by repressing or inducing its synthesis,
or controlling the amounts of other enzymes which degrade it. However the
activity of individual enzyme molecules may be reduced by inhibitors or
increased by effectors.
An enzyme molecule will be inactivated if the substrate can no longer bind to
the active site. This may be effected by an inhibitor covalently bonding to
the site, or binding very tightly so that its dissociation is very slow. This
is irreversible inhibition (see the example of chymotrypsin below).
In reversible inhibition, there is a rapid equilibrium of the enzyme and
inhibitor. Competitive inhibitors bind to the active site.
Non-competitive inhibitors bind elsewhere, but reduce the rate constant
(the "turnover number") of the formation of enzyme E and product P from the
ES complex. Effector molecules may act by having an opposite effect.
In oligomeric enzymes with several active sites, allosteric inhibitors and
effectors associating with one site affect the binding capabilities of
others (see above on allosteric interactions).
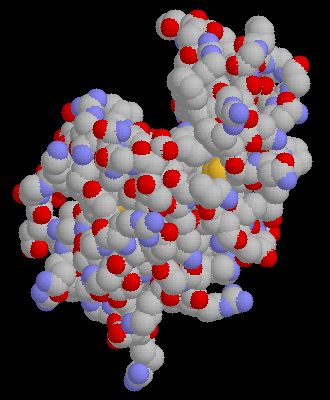
 Click here for a
space-filling model of potato carboxypeptidase A inhibitor (38 residues long)
binding to the active site of bovine carboxypeptidase A.
Click here for a
space-filling model of potato carboxypeptidase A inhibitor (38 residues long)
binding to the active site of bovine carboxypeptidase A.
 Examine the pdb file with RasMol.
4cpa (235Kb)
[Bbk|BNL|ExP|Waw|Hal]
Examine the pdb file with RasMol.
4cpa (235Kb)
[Bbk|BNL|ExP|Waw|Hal]
Coenzymes
Many enzymes require coenzymes in order to function. Coenzymes are bound
at the active site in order to interact with the substrate and play an essential
role in the catalysed reaction. They act as carriers of a variety of chemical
groups. Most water-soluble vitamins are components of coenzymes. The complex
of enzyme + coenzyme is called the holoenzyme. The enzyme minus its coenzyme is
referred to as the apoenzyme.
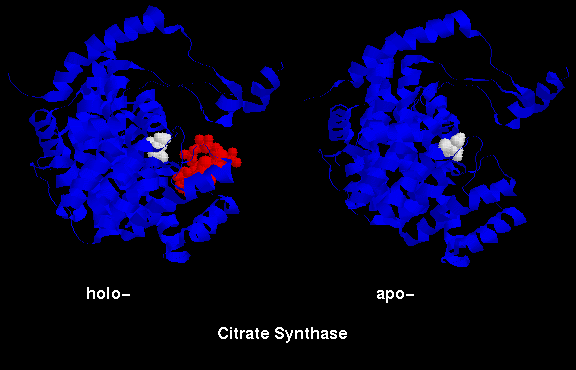
An example of this is citrate synthase (or "synthetase").
In one form,
the surrounding domains are closed around the active site with both the product
(citrate) and coenzyme (coenzyme A, or CoA) bound. In the open
form, only citrate is bound (see below).
- closed form (holoenzyme), showing both citrate and CoA. 2cts (313Kb)
[Bbk|BNL|ExP|Waw|Hal]
- open form (apoenzyme), with citrate only 1cts (293Kb)
[Bbk|BNL|ExP|Waw|Hal]
24 Kb GIF
This RasMol SCRIPT highlights the citrate ion (white),
and, in the case of the holoenzyme, CoA (red).
The function of the enzyme is to catalyze one of the reactions of the citric acid
cycle, in which an acetyl group, covalently bound to CoA (in the form of
acetyl-CoA) reacts with the substrate and 1 water moleculeto give citrate
and CoA. CoA is a universal carrier of acyl (in many cases acetyl) groups. Acetyl-CoA
has a high acetyl group transfer potential.
Click here for a diagram (courtesy of SWISS-3DIMAGE, maintained by Manuel Peitsch ) of the closed form of citrate synthase, indicating
the structure of CoA (it consists of an ADP 3'-phosphate, a pantothenate
and a 2-aminoethanethiol group, covalently bonded), and also including the
citrate ion.
NAD+ in dehydrogenases
The coenzyme NAD+ (nicotinamide adenine dinucleotide) is involved in the function
of certain dehydrogenases. The crystal structures of various enzymes of this
type - listed below - reveal that each consist of 2
domains, one of which, the NAD+ binding domain, is similar in each case. The
other domain differs markedly; although (1) and (2) are very similar and appear
to have evolved from a common precursor. Note that in some of these crystal
structures, there is more than one enzyme molecule in the asymmetric group.
In each case, selecting the Colour:chain option on RasMol highlights the
bound nucleotide. Typing select NAD followed by spacefill will
also help.
- dogfish lactate dehydrogenase 3ldh (241Kb) [Bbk|BNL|ExP|Waw|Hal],
- soluble porcine malate dehydrogenase 4mdh (507Kb) [Bbk|BNL|ExP|Waw|Hal],
- horse liveralcohol dehydrogenase 6adh (503Kb) [Bbk|BNL|ExP|Waw|Hal],
- lobster glyceraldehyde-3-phosphate dehydrogenase 1gpd (448Kb) [Bbk|BNL|ExP|Waw|Hal],
- Bacillus stearothermophilus glyceraldehyde-3-phosphate dehydrogenase 1gd1 (966Kb) [Bbk|BNL|ExP|Waw|Hal]
SWISS-3DIMAGE provides
two
different
views
of the Bacillus stearothermophilus apoenzyme (5), and also
several images of horse liver alcohol dehydrogenase:
-
firstly, highlighting the bound NAD and the zinc ion at the bottom of a hydrophobic
pocket formed at the junction of the catalytic and nucleotide-binding domains;
this structure is also complexed with a bound dimethyl sulphoxide (DMSO)
molecule
-
secondly, a more detailed view of the above indicating the residues which
bind to these groups;
-
three
different
"close-up" views (in stereo) of these groups in the binding site.
The NAD+ binding domain is also present in other nucleotide-
binding proteins, but also occurs in some proteins, eg flavodoxin, which do
not bind nucleotides. Possibly this is simply a result of the finite number of ways
in which a polypeptide chain may be folded, rather than being the product of
divergent evolution.
flavodoxin 1flv (127Kb) [Bbk|BNL|ExP|Waw|Hal]
Index to Course Material
Index to Section 12
Enzymes Index;
 Part 2
Part 2
Last updated 11th Jul '96
 Click here for a
space-filling model of potato carboxypeptidase A inhibitor (38 residues long)
binding to the active site of bovine carboxypeptidase A.
Click here for a
space-filling model of potato carboxypeptidase A inhibitor (38 residues long)
binding to the active site of bovine carboxypeptidase A.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}